Экологическая толерантность: Экологическая толерантность
12. Экологическая толерантность животных
Толерантность
или экологическая валентность – это
степень, в которой организмы способны
переносить колебания факторов.
Фактор
среды, ограничивающий возможности
существования и распространения
организмов, называется лимитирующим.
По удачному выражению известного эколога
Ю. Одума, организмы не являются рабами
физических воздействий. Они приспосабливаются
сами и изменяют условия среды так, чтобы
ослабить лимитирующее влияние температуры,
света, влажности и т. д. Подобная
компенсация факторов возможна большей
частью на уровне не отдельной особи, а
сообщества и популяций вида. Выражается
она существованием различных популяций
— экотипов — у вида с широким географическим
распространением, оптимумы и пределы
толерантности которых соответствуют
условиям конкретных районов и их
отдельных участков. Например,
медуза-сердцевидка Aurelia aurita имеет
несколько экотипов, приспособленных к
различным температурным условиям:
обитатели северных районов активно
плавают при температурах, которые южные
популяции привели бы к полному подавлению.
Многощетинковый червь нереида Nereis
diversicolor живет в Черном море и в замкнутых
лагунах на побережье Румынии. Если
лагунную популяцию перенести в Черное
море, она погибнет, так как там иное
содержание солей. Такой же эффект
наблюдается при перенесении черноморской
популяции в лагуны. Между тем эти
популяции представляют собой лишь
биологические расы одного и того же
вида.
Для
распознавания лимитирующих факторов
необходимо изучать виды на границах их
ареалов. Без этого часто трудно сказать,
что же именно не позволяет виду расширить
занимаемую им область. Так, после 1880 г.
в Финляндии наблюдалось незначительное
повышение средних температур. Одновременно
было установлено расширение на север
ареала чибиса Vanellus vanellus. Это говорит о
том, что именно температура является
лимитирующим фактором, поскольку
остальные условия оставались в тот
период неизменными.
Более
подробно отношения животных организмов
к конкрет-ным факторам среды будут
рассмотрены ниже, причем применительно
к основным биоциклам — морскому,
пресноводному и сухопутному.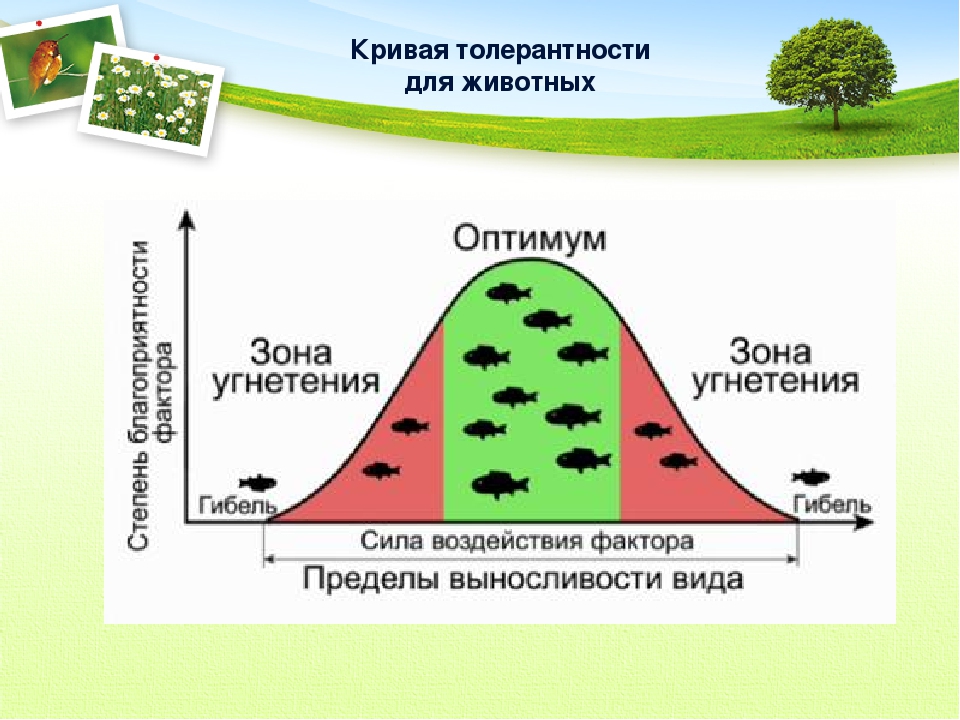
13.Информация как экологический фактор
ЭКОЛОГИЧЕСКИЙ
ФАКТОР — любое условие среды, на которое
живое реагирует приспособительными
реакциями.
Экологический
фактор — это любой элемент среды,
оказывающий прямое или косвенное влияние
на живые организмы хотя бы на протяжении
одной из фаз их развития.
По своей
природе экологические факторы делят,
по крайней мере, на три группы:
абиотические
факторы — влияния неживой природы;
биотические
факторы — влияния живой природы.
антропогенные
факторы — влияния, вызванные разумной
и неразумной деятельностью человека
(«антропос» — человек).Человек
видоизменяет живую и неживую природу,
и берет на себя в известном смысле и
геохимическую роль (например, высвобождая
замурованный в виде угля и нефти на
многие миллионы лет углерод и выпуская
его в воздух углекислым газом). Поэтому
антропогенные факторы по размаху и
глобальности своего воздействия
приближаются к геологическим силам.Не
редко экологические факторы подвергают
и более детальной классификации, когда
надо указать на какую-то конкретную
группу факторов.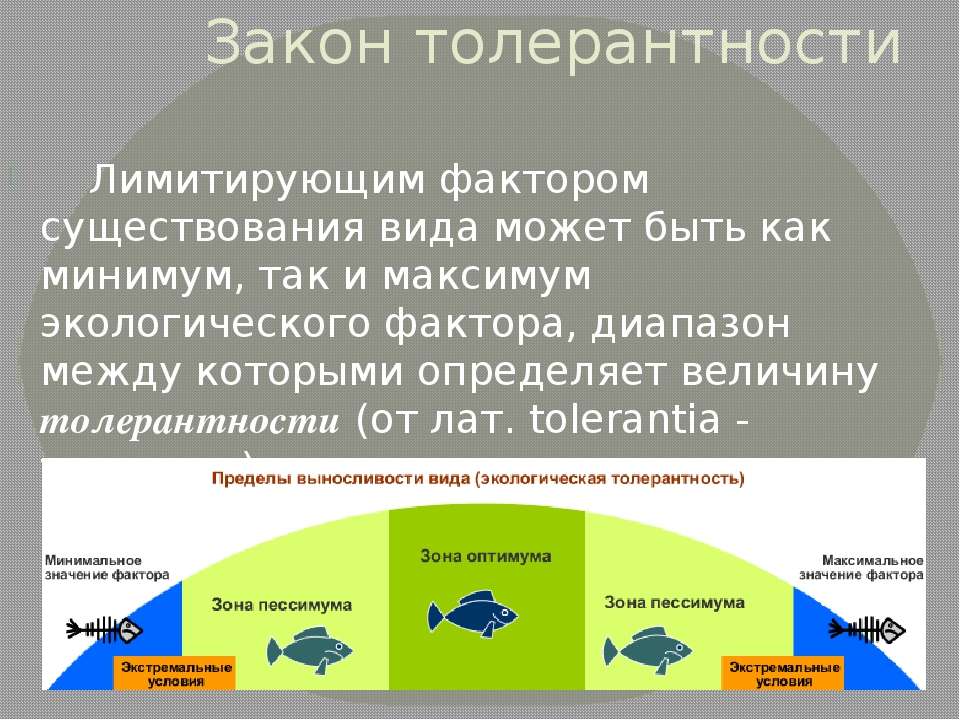 Например, различают
Например, различают
климатические (относящиеся к климату),
эдафические (почвенные) факторы среды.В
качестве хрестоматийного примера
опосредованного действия экологических
факторов приводят так называемые птичьи
базары, представляющие собой огромные
скопления птиц. Высокая плотность птиц
объясняется целой цепочкой
причинно-следственных связей. Птичий
помет попадает в воду, органические
вещества в воде минерализуются бактериями,
повышенная концентрация минеральных
веществ приводит к повышению численности
водорослей, а вслед за ними — и зоопланктона.
Низшими ракообразными, входящими в
зоопланктон, питаются рыбы, а рыбами —
птицы, населяющие птичий базар. Цепочка
замыкается. Птичий помет выступает в
качестве экологического фактора,
опосредованно повышающего численность
колонии птиц.Как же сопоставлять действие
столь разных по природе факторов? Не
смотря на огромное множество факторов,
из самого определения экологического
фактора как элемента среды, оказывающего
влияние на организм, следует нечто
общее.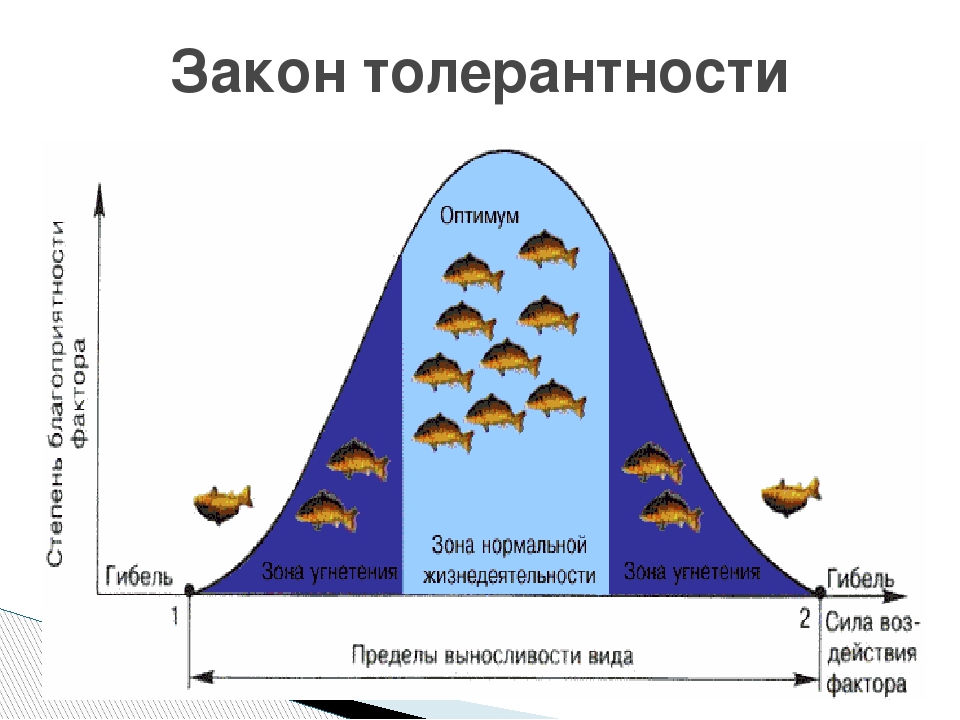 А именно: действие экологических
А именно: действие экологических
факторов всегда выражается в изменении
жизнедеятельности организмов, а в
конечном итоге, — приводит к изменению
численности популяции. Это и позволяет
сравнивать действие различных
экологических факторов.Стоит ли говорить,
что действие фактора на особь определяется
не природой фактора, а его дозой . В свете
сказанного выше, да и простого жизненного
опыта, становится очевидным, что эффект
определяет именно доза фактора.
Действительно, что такое фактор
«температура»? Это в достаточной
степени абстракция, а вот если сказать,
что температура -40 по Цельсию — тут уже
не до абстракций, поскорее бы закутаться
во все теплое! С другой стороны, +50
градусов нам покажутся не многим
лучше.Таким образом, фактор воздействует
на организм определенной дозой, и среди
этих доз можно выделить минимальные,
максимальные и оптимальные дозы, а также
те значения, при которых жизнь особи
прекращается (их называют летальными,
или смертельными).
Толерантность — Экологические термины и определения
Способность организма выносить отелонения экологических факторов среды от оптимальных для него значений.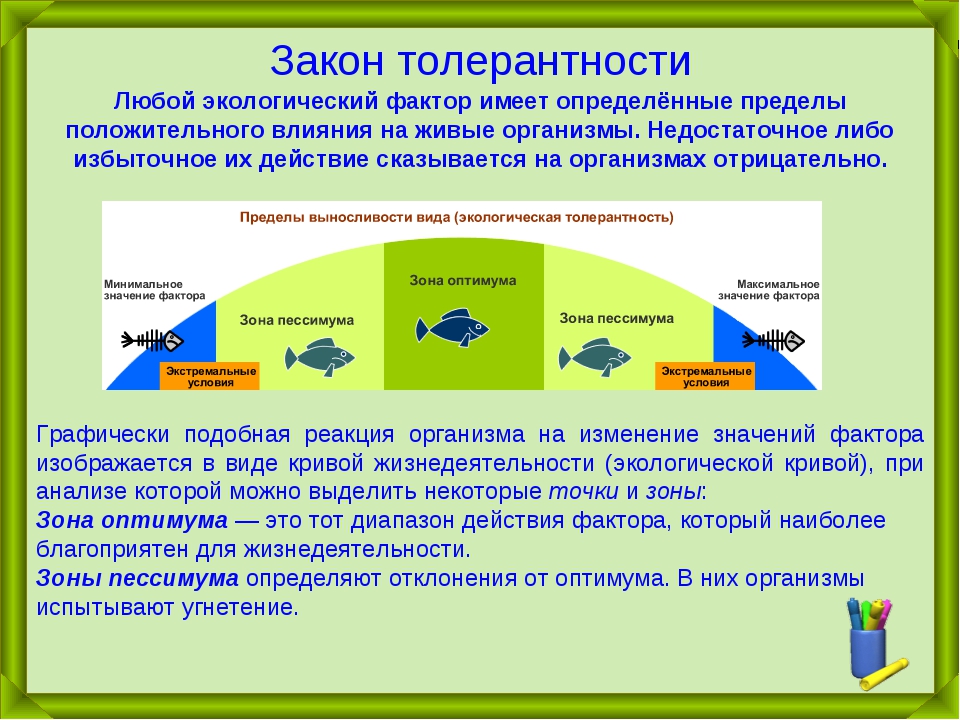 Организмы с широким диапазоном толерантности обозначаются приставкой «эври-«, а с узким диапазоном толерантности — приставкой «стено-» (эврибионт, стенобионт).
Организмы с широким диапазоном толерантности обозначаются приставкой «эври-«, а с узким диапазоном толерантности — приставкой «стено-» (эврибионт, стенобионт).
Источник:
Словарь экологических терминов и определений
на Gufo.me
Значения в других словарях
- Толерантность —
(от лат. tolerantia — терпение) иммунологическая, отсутствие или ослабление иммунологического ответа на данный антиген (См. Антигены) при сохранении иммунореактивности ко всем прочим антигенам. Термин введён в 1953 английским иммунологом…
Большая советская энциклопедия - толерантность —
ТОЛЕРАНТНОСТЬ иммунологическая (от лат. tolerantia — терпение), иммунологич. феномен, характеризующийся ареактивностью клеток организма к определённым антигенам.
tolerantia — терпение), иммунологич. феномен, характеризующийся ареактивностью клеток организма к определённым антигенам.
Ветеринарный энциклопедический словарь - Толерантность —
(лат. tolerantia – способность переносить). В психофармакологии: способность переносить воздействие лечебных доз лекарств без заметного терапевтического или токсического эффекта. В более широком смысле говорят о терапевтической Т. в неподдающихся лечению случаях психических заболеваний.
Толковый словарь психиатрических терминов - толерантность —
Толерантность, толерантности, толерантности, толерантностей, толерантности, толерантностям, толерантность, толерантности, толерантностью, толерантностями, толерантности, толерантностях
Грамматический словарь Зализняка - толерантность —
[< лат. tolerantia терпение] – терпимость к чужим мнениям и верованиям
Большой словарь иностранных слов - толерантность —
толерантность I ж. Отсутствие или ослабление реакции на какой-либо неблагоприятный фактор, объясняющееся снижением чувствительности к его воздействию. II ж. Снисходительная терпимость к кому-либо или к чему-либо (чужому мнению, иному верованию и т.п.
Толковый словарь Ефремовой - толерантность —
Толерантность — устойчивость живых организмов к действию факторов внешней среды: высокой температуре (термо…), повышенной концентрации солей (гало…), углеводов (осмо…) и т. д.
Толковый словарь по почвоведению - толерантность —
ТОЛЕРАНТНОСТЬ Свойство личности, проявляющееся в терпимости к различным мнениям, внешним воздействиям, в выносливости по отношению к неблагоприятным эмоциональным факторам, в т. ч. связанным с соревновательной борьбой. (Терминология спорта. Толковый словарь спортивных терминов, 2001)
Словарь спортивных терминов - толерантность —
сущ., кол-во синонимов: 12 либерализм 17 либеральность 16 мягкость 52 невзыскательность 17 нетребовательность 15 политкорректность 2 привыкание 10 снисходительное отношение 5 снисходительность 17 снисхождение 15 терпение 13 терпимость 21
Словарь синонимов русского языка - ТОЛЕРАНТНОСТЬ —
ТОЛЕРАНТНОСТЬ (от лат. toleran-tia — терпение) — англ. tolerance; toleration; нем. Toleranz. 1. Терпимость к чужому образу жизни, поведению, обычаям, чувствам, мнениям, идеям, верованиям.
Социологический словарь - Толерантность —
(лат. tolerantia способность переносить, терпеливость; син. переносимость) в фармакологии — способность организма переносить воздействие определенного лекарственного вещества или яда без развития соответствующего терапевтического или токсического эффекта.
Медицинская энциклопедия - Толерантность —
(в физиологической антропологии) (лат. tolerantia терпение) — «терпимость» иммунной системы организма (отсутствие или ослабление иммунологического ответа) к конкретным антигенам.
Физическая антропология - толерантность —
ТОЛЕРАНТНОСТЬ и, ж. tolérant adj. 1. устар. Терпимое, снисходительное отношение к кому-, чему-л. БАС-1. Терпимость вообще и особенно относительно религиозных мнений, короче — веротерпимость. Павленков 1911.
Словарь галлицизмов русского языка - ТОЛЕРАНТНОСТЬ —
ТОЛЕРАНТНОСТЬ (от лат. tolerantia – терпимость) – качество, характеризующее отношение к другому человеку как к равнодостойной личности и выражающееся в сознательном подавлении чувства неприятия, вызванного всем тем…
Новая философская энциклопедия - толерантность —
орф. толерантность, -и
Орфографический словарь Лопатина - Толерантность —
Бинарное отношение на множестве А, обладающее свойствами рефлексивности и симметричности, т. е. удовлетворяющее условиям aRa для всех и aRb влечет за собой bRa для любых Т. Rна универсальной алгебре наз.
Математическая энциклопедия - толерантность —
См.: языковая терпимость.
Словарь лингвистических терминов Жеребило - толерантность —
толерантность , -и
Орфографический словарь. Одно Н или два? - Толерантность —
(от лат. tolerantia — терпение) терпимость к чужому образу жизни, мнению, поведению. (Иванова О.А. Конфликты в школьной среде. — СПб., 2003. С. 184) одно из важнейших профессиональных качеств учителя. В основе…
Педагогический терминологический словарь - толерантность —
ТОЛЕР’АНТНОСТЬ, толерантности, мн. нет, ·жен. (·книж. ). ·отвлеч. сущ. к толерантный; терпимость.
Толковый словарь Ушакова - Толерантность —
(лат. tolerantia – терпимость) – термин, обозначающий терпимость к чужим мнениям, верованиям, поведению.
Словарь по культурологии - ТОЛЕРАНТНОСТЬ —
ТОЛЕРАНТНОСТЬ (от лат. tolerantia — терпение) — .. 1) иммунологическое состояние организма, при котором он неспособен синтезировать антитела в ответ на введение определенного антигена при сохранении иммунной реактивности к другим антигенам.
Большой энциклопедический словарь - толерантность —
(от лат. tolerantia — терпение) иммунологическая, отсутствие или ослабление иммунологич. ответа на данный антиген при сохранении иммунореактивности организма ко всем прочим антигенам. Термин введён в 1953…
Биологический энциклопедический словарь - толерантность —
ТОЛЕРАНТНОСТЬ — устойчивость живых организмов к действию факторов внешней среды: высокой температуре (термотолерантность), повышенной концентрации солей (галотолерантность), углеводов (осмотолерантность) и т. д.
Ботаника. Словарь терминов - толерантность —
Толера́нтн/ость/.
Морфемно-орфографический словарь - толерантность —
Терпимость Ср. Я не могу относиться к нему толерантно, когда он утверждает, что правительство нарочно спаивает народ водкой. Достоевский. Бесы. 2, 4, 3. Ср.
Фразеологический словарь Михельсона - Толерантность —
См. Языковая терпимость
Словарь социолингвистических терминов
 tolerantia — терпение), иммунологич. феномен, характеризующийся ареактивностью клеток организма к определённым антигенам.
tolerantia — терпение), иммунологич. феномен, характеризующийся ареактивностью клеток организма к определённым антигенам. tolerantia терпение] – терпимость к чужим мнениям и верованиям
tolerantia терпение] – терпимость к чужим мнениям и верованиям ч. связанным с соревновательной борьбой. (Терминология спорта. Толковый словарь спортивных терминов, 2001)
ч. связанным с соревновательной борьбой. (Терминология спорта. Толковый словарь спортивных терминов, 2001)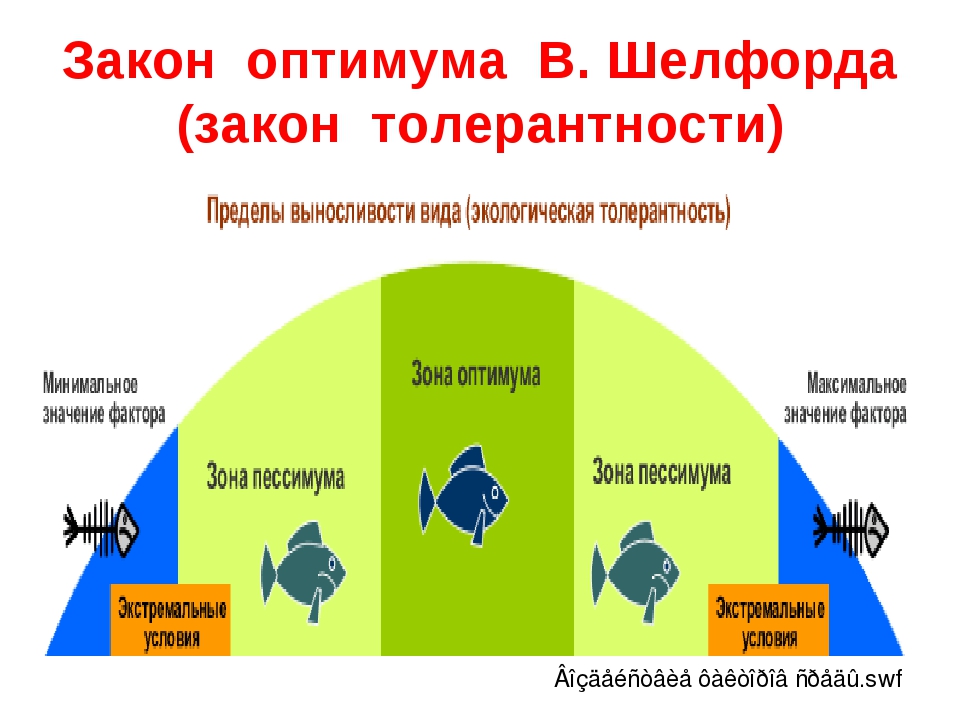 tolerantia способность переносить, терпеливость; син. переносимость) в фармакологии — способность организма переносить воздействие определенного лекарственного вещества или яда без развития соответствующего терапевтического или токсического эффекта.
tolerantia способность переносить, терпеливость; син. переносимость) в фармакологии — способность организма переносить воздействие определенного лекарственного вещества или яда без развития соответствующего терапевтического или токсического эффекта. Павленков 1911.
Павленков 1911. е. удовлетворяющее условиям aRa для всех и aRb влечет за собой bRa для любых Т. Rна универсальной алгебре наз.
е. удовлетворяющее условиям aRa для всех и aRb влечет за собой bRa для любых Т. Rна универсальной алгебре наз.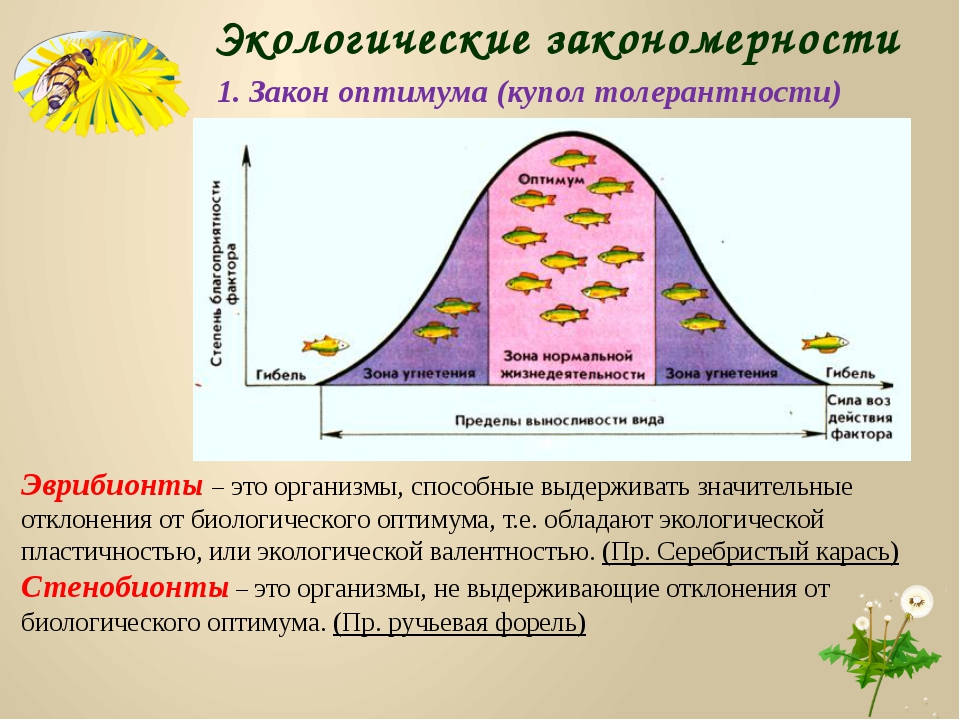 В основе…
В основе…

Экологическая пластичность флоры урочища «Лысая гора» (Студеный овраг, Красноглинский район г. о. Самара)
Авторы:
Ильина Валентина Николаевна
Поволжская государственная социально-гуманитарная академия (Самара)
Журнал: Научный диалог
Номер: 3(15)
Год: 2013
Страницы: 43-56
DOI:
УДК:
Ключевые слова
флора Лысая гора экологическая пластичность экологическая толерантность стенобионты flora “Lysaya Gora” ecological plasticity ecological tolerance stenobionts
Аннотация
В статье рассматривается видовой состав флоры естественных лесостепных урочищ в черте крупных городов на примере Лысой горы (г. Самара), испытывающих сильную антропогенную нагрузку, а также принадлежность видов растений урочища к группам экологической пластичности (толерантности).
Самара), испытывающих сильную антропогенную нагрузку, а также принадлежность видов растений урочища к группам экологической пластичности (толерантности).
Источники
Жукова Л. А. Экологические шкалы и методы анализа экологического разнообразия растений : монография / Л. А. Жукова [и др.]. – Йошкар-Ола : Изд-во МарГУ, 2010. – 368 с.
Ильина В. Н. О современном состоянии растительного покрова горы Тип-Тяв (Сокольи горы, Самарская обл.) / В. Н. Ильина // Вопросы степеведения. – Оренбург : Институт степи УрО РАН, 2010. – С. 26–33.
Ильина В. Н. Экологическая пластичность видов флоры урочища «Верховья реки Бинарадки» / В. Н. Ильина // Репродуктивная биология, география и экология растений и сообществ Среднего Поволжья : Материалы Всеросс. конф. (27–29 ноября 2012 г.). – Ульяновск : Изд-во УлГПУ, 2012. – С. 107–109.
Ильина Н. С. Динамика растительного покрова лесостепи под влиянием антропогенных факторов / Н. С. Ильина [и др.] // Взаимодействие человека и природы на границе Европы и Азии : тез. докл. научно-практ. конфер., Самара, 18–20 дек., 1996 ; [отв. ред. В. И. Матвеев, И. Б. Васильев]. – Самара : Ком. экологии и природ. ресурсов Самар. обл. : Ин-т истории и археологии Поволжья, 1996. – С. 68–71.
С. Ильина [и др.] // Взаимодействие человека и природы на границе Европы и Азии : тез. докл. научно-практ. конфер., Самара, 18–20 дек., 1996 ; [отв. ред. В. И. Матвеев, И. Б. Васильев]. – Самара : Ком. экологии и природ. ресурсов Самар. обл. : Ин-т истории и археологии Поволжья, 1996. – С. 68–71.
Ильминских Н. Г. Анализ городской флоры (на примере флоры города Казани) : автореферат диссертации… кандидата биологических наук / Н. Г. Ильминских. – Санкт-Петербург, 1982. – 20 с.
Кавеленова Л. М. Некоторые проблемы изучения городской растительности / Л. М. Кавеленова, А. Г. Здетоветский // Экологическая безопасность городов : проблемы решения на муниципальном уровне : материалы Всероссийской научно-практической конференции 16–19 мая 2000 г. – Самара : [б. и.], 2000. – С. 59–60.
Кавеленова Л. М. Проблемы организации системы фитомониторинга городской среды в условиях лесостепи / Л. М. Кавеленова. – Самара : Изд-во Самарского университета, 2003. – 124 с.
Матвеева Н. В. К изучению флоры города Самары / Н. В. Матвеева // Региональные экологические проблемы и возможные пути их реализации : тезисы докладов межвузовской научно-практической конференции студентов и молодых специалистов, Самара, 21–22 декабря 1994 г. – Самара : Изд-во СГПУ, 1994. – С. 19.
В. К изучению флоры города Самары / Н. В. Матвеева // Региональные экологические проблемы и возможные пути их реализации : тезисы докладов межвузовской научно-практической конференции студентов и молодых специалистов, Самара, 21–22 декабря 1994 г. – Самара : Изд-во СГПУ, 1994. – С. 19.
Матвеева Н. В. Некоторые закономерности в распространении флоры на территории города Самары / Н. В. Матвеева // Взаимодействие человека и природы на границе Европы и Азии : тезисы докладов конференции, Самара, 18–20 декабря 1996 г. – Самара : [б. и.], 1996. – С. 104–105.
Мерзлякова И. Е. Итоги изучения флоры г. Томска / И. Е. Мерзлякова // Сравнительная характеристика на рубеже III тысячелетия : достижения, проблемы, перспективы : материалы V рабочего совещания по сравнительной флористике. – Санкт-Петербург : БИН РАН, 2000. – С. 256–259.
Мозговая О. А. Городская флора – показатель устойчивости растений к воздействию урбанизации / О. А. Мозговая // Актуальные вопросы ботаники в СССР : Тезисы докладов 8 делегатского съезда Всесоюзного ботанического общества.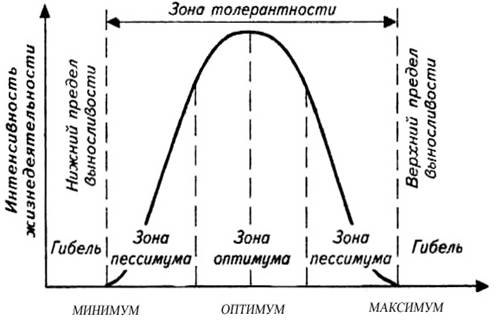 – Алма-Ата : [б. и.], 1988. – С. 427–428.
– Алма-Ата : [б. и.], 1988. – С. 427–428.
Плаксина Т. И. Конспект флоры Волго-Уральского региона / Т. И. Плаксина. – Самара : Самарский университет, 2001. – 388 с.
Прохоров В. Е. Многолетняя динамика флоры города Казани / В. Е. Прохоров, В. В. Старцева // Окружающая среда и устойчивое развитие регионов : новые методы и технологии исследований : К 70-летию географического и 20-летию экологического факультетов КГУ : труды Всеросс. науч. конф. с международным участием:– Казань : Бриг, 2009. – Том III : Моделирование в охране окружающей среды. Общая экология и охрана биоразнообразия. – С. 264–268.
Раков Н. С. Флора города Ульяновска и его окрестностей / Н. С. Раков. – Ульяновск : Корпорация технологий продвижения, 2003. – 216 с.
Серебряков И. Г. Жизненные формы высших растений и их изучение / И. Г. Серебряков // Полевая геоботаника. – Т. 3. – Москва ; Ленинград : Наука, 1964. – С. 146–205.
Сосудистые растения Самарской области : учебное пособие. – Самара : ИПК Содружество, 2007. – 400 с.
– 400 с.
Флора Самарской области : учебное пособие. – Самара : Изд-во СГПУ, 2007. – 321 с.
Цыганов Д. Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов / Д. Н. Цыганов. – Москва : Наука, 1983. – 196 с.
Щербиновский Н. Дневники Самарской природы 1916 года / Н. Щербиновский. – Самара : Типография Губернского Совета Народного хозяйства, 1919. – № 2. – 146 с.
Полный текст статьи
скачать
Экологическая пластичность флоры урочища «Лысая гора» (Студеный овраг, Красноглинский район г. о. Самара)
Please use this identifier to cite or link to this item:
https://elar.rsvpu.ru/handle/123456789/19773
| Title: | Экологическая пластичность флоры урочища «Лысая гора» (Студеный овраг, Красноглинский район г.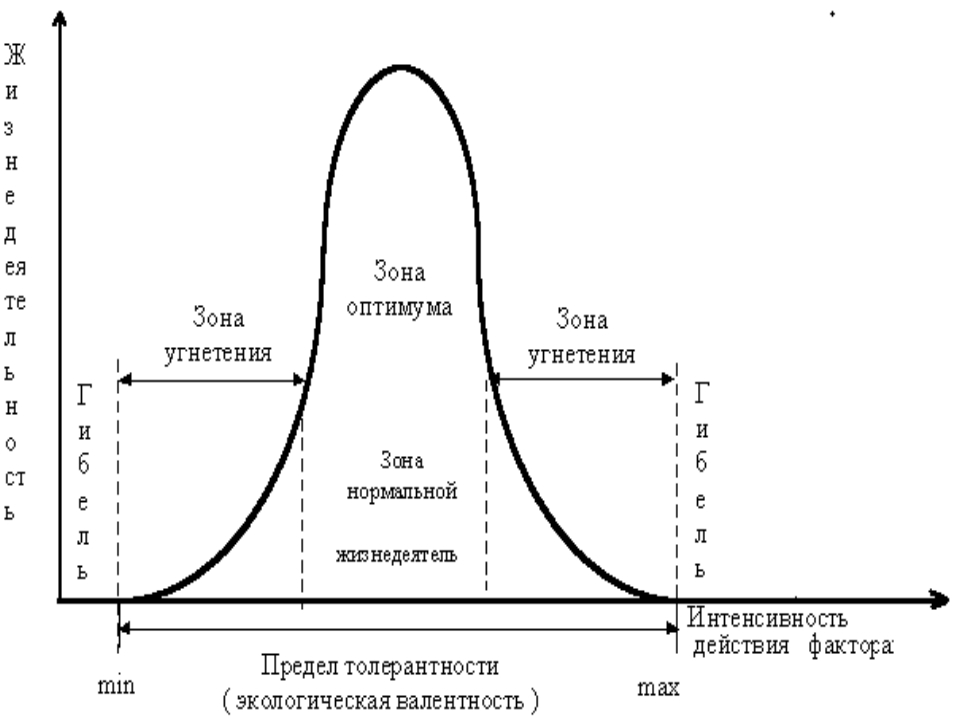 о. Самара) о. Самара) |
| Other Titles: | Ecological Plasticity of Flora in «Lysaya Gora» Stow (Studenyi Ovrag, Krasnoglinsky City District, City of Samara) |
| Authors: | Ilyina, V. N. Ильина, В. Н. |
| Issue Date: | 2013 |
| Publisher: | Российский государственный профессионально-педагогический университет |
| Citation: | Ильина, В. Н. Экологическая пластичность флоры урочища «Лысая гора» (Студеный овраг, Красноглинский район г. о. Самара) / В. Н. Ильина // Научный диалог. — 2013. — № 3(15) : Естествознание. Экология. Науки о земле. — С. 43-56. |
| Abstract: | The article studies the plant species composition in natural forest-steppe stows within the boundaries of big cities by the example of «Lysaya Gora» (Samara) which is under a heavy anthropogenic load as well as the stow’s plant species belonging to the groups of ecological plasticity (tolerance) В статье рассматривается видовой состав флоры естественных лесостепных урочищ в черте крупных городов на примере Лысой горы (г.  Самара), испытывающих сильную антропогенную нагрузку, а также принадлежность видов растений урочища к группам экологической пластичности (толерантности) Самара), испытывающих сильную антропогенную нагрузку, а также принадлежность видов растений урочища к группам экологической пластичности (толерантности) |
| Keywords: | ФЛОРА FLORA ЛЫСАЯ ГОРА ЭКОЛОГИЧЕСКАЯ ПЛАСТИЧНОСТЬ ECOLOGICAL PLASTICITY ЭКОЛОГИЧЕСКАЯ ТОЛЕРАНТНОСТЬ ECOLOGICAL TOLERANCE СТЕНОБИОНТЫ LYSAYA GORA STENOBIONTS |
| ISSN: | 2225-756X |
| Origin: | Научный диалог. — 2013. — № 15 |
| Appears in Collections: | Научный диалог |
Items in DSpace are protected by copyright, with all rights reserved, unless otherwise indicated.
Экологическая толерантность животных — КиберПедия
Факторы среды изменчивы. Примером могут служить колебания температуры воздуха в течение суток и года или изменения ее при перемещении от экватора к полюсам. Нормальное существование животных наблюдается только при тех изменениях экологического фактора, которые находятся в пределах его минимального и максимального значений. Эти значения представляют собой границы толерантности (пределы выносливости) относительно данного фактора, его экологический диапазон (рис. 111).
Эти значения представляют собой границы толерантности (пределы выносливости) относительно данного фактора, его экологический диапазон (рис. 111).
Рис. 111. Границы толерантности организмов в зависимости от
интенсивности экологического фактора (из Р.Дажо, 1975)
Между минимумом и максимумом, ограничивающими возможность существования животного, находится относительно узкая зона оптимума, наиболее для него благоприятная. Между зоной оптимума и границей, определяющей возможность существования вида, лежит зона пессимума, где организм оказывается в плохих условиях. Зона пессимума, как и зона оптимума, устанавливается по плотности популяций вида, т. е. по числу особей, приходящихся на единицу площади. Конечно, в зоне оптимума вид будет представлен более плотными популяциями.
По аналогии с химической валентностью экологи ввели понятие «экологическая валентность». Экологическая валентность вида — это его способность заселять различную среду, характеризующуюся определенными изменениями экологических факторов. Вид с низкой экологической валентностью, способный переносить лишь ограниченные колебания факторов среды, называют стенобионтным. Примерами стенобионтных видов сухопутных животных являются человекообразная обезьяна шимпанзе, обитающая в тропических влажных лесах, или наш глухарь — житель хвойного леса умеренной зоны. В противоположность этому вид, способный заселять различные местообитания или местообитания с изменчивыми условиями, называют эврибионтным. Примером эврибионтного вида может служить лисица, встречающаяся на равнине и в горах, в лесу и в степи, в пределах нескольких ландшафтно-климатических зон. К типичным стенобионтам, обитающим в водной среде, относятся рифообразующие кораллы, требующие строго определенных (высокой) температуры, солености и степени освещенности морской воды. Они распространены в пределах тропической зоны Мирового океана. Киты же, будучи эврибионтами, при наличии пищи живут в различных широтах.
Вид с низкой экологической валентностью, способный переносить лишь ограниченные колебания факторов среды, называют стенобионтным. Примерами стенобионтных видов сухопутных животных являются человекообразная обезьяна шимпанзе, обитающая в тропических влажных лесах, или наш глухарь — житель хвойного леса умеренной зоны. В противоположность этому вид, способный заселять различные местообитания или местообитания с изменчивыми условиями, называют эврибионтным. Примером эврибионтного вида может служить лисица, встречающаяся на равнине и в горах, в лесу и в степи, в пределах нескольких ландшафтно-климатических зон. К типичным стенобионтам, обитающим в водной среде, относятся рифообразующие кораллы, требующие строго определенных (высокой) температуры, солености и степени освещенности морской воды. Они распространены в пределах тропической зоны Мирового океана. Киты же, будучи эврибионтами, при наличии пищи живут в различных широтах.
Для выражения относительной степени толерантности употребляют соответствующие данным факторам термины с добавлением приставки «стено» (узкий) или «эври» (широкий). Так, относительно температуры животные бывают стенотермными или эвритермными, солености водной среды — стеногалинными или эвригалинными, пищевого рациона — стенофагами или эврифагами и т. д. Наконец, в отношении местообитания их можно разделить на стеноэков и эвриэков.
Так, относительно температуры животные бывают стенотермными или эвритермными, солености водной среды — стеногалинными или эвригалинными, пищевого рациона — стенофагами или эврифагами и т. д. Наконец, в отношении местообитания их можно разделить на стеноэков и эвриэков.
Фактор среды, ограничивающий возможности существования и распространения организмов, называется лимитирующим. Для распознавания лимитирующих факторов необходимо изучать виды на границах их ареалов.
Условия существования и распространения
Животных в море
Океаны и моря представляют собой самый крупный биоцикл Земли. На их долю выпадает 71 % поверхности планеты. В то же время они включают и наиболее богатую фауну, которая составляет 64 % видов животных, тогда как на сушу приходится только 36 %. Это и понятно, ведь жизнь зародилась в море, до настоящего времени здесь обитают представители очень многих классов животных, за исключением огромного большинства насекомых, многоножек и амфибий. Многие классы животных обитают только в море. К ним относятся коралловые полипы, плеченогие, боко-нервные и головоногие моллюски, бесчерепные, оболочники, губки, многощетинковые кольчецы, немертины и др. Вместе с тем следует отметить, что в океане и поныне есть представители очень древних групп животных, которые за миллионы лет изменились сравнительно мало. Это свидетельствует о замедленности темпов эволюции морских животных по сравнению с наземными.
Многие классы животных обитают только в море. К ним относятся коралловые полипы, плеченогие, боко-нервные и головоногие моллюски, бесчерепные, оболочники, губки, многощетинковые кольчецы, немертины и др. Вместе с тем следует отметить, что в океане и поныне есть представители очень древних групп животных, которые за миллионы лет изменились сравнительно мало. Это свидетельствует о замедленности темпов эволюции морских животных по сравнению с наземными.
Между наземными и водными средами наблюдаются существенные различия в их физико-химических свойствах. Особое экологическое значение для морских организмов имеют плотность, давление, глубина проникновения солнечной радиации, распределение тепла, содержание газов и растворенных солей, течения.
Среди морских животных по их отношению к давлению различают эврибатные и стенобатные виды.
На распределение животных в море сильное влияние оказывает свет, вернее, степень проникновения солнечной радиации, которая зависит от веществ, растворенных и взвешенных в воде.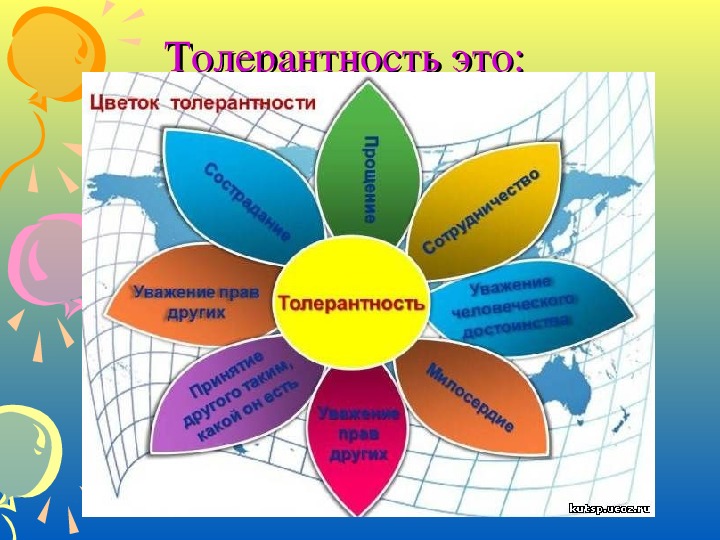 По мере увеличения глубины степень проникновения солнечной радиации уменьшается, причем довольно быстро. На глубине 1 м инфракрасное излучение полностью поглощается, а видимый свет наполовину слабее, чем на поверхности. На глубине 200-400 м света уже не хватает для существования растений. Большие глубины практически лишены освещения, и животные там обитают в темноте. Толщу воды в море принято делить на зоны: хорошо освещенную эвфотическую (от 0 до 30 м), дисфотическую (30— 200 м) и лишенную света афотическую (ниже 200 м).
По мере увеличения глубины степень проникновения солнечной радиации уменьшается, причем довольно быстро. На глубине 1 м инфракрасное излучение полностью поглощается, а видимый свет наполовину слабее, чем на поверхности. На глубине 200-400 м света уже не хватает для существования растений. Большие глубины практически лишены освещения, и животные там обитают в темноте. Толщу воды в море принято делить на зоны: хорошо освещенную эвфотическую (от 0 до 30 м), дисфотическую (30— 200 м) и лишенную света афотическую (ниже 200 м).
Важную роль в жизни водных животных играет распределение в морях тепла. Источником его служит энергия солнечной радиации, поэтому распределение температур на поверхности и в верхнем слое воды зависит от климата соответствующей части земного шара, где располагается данный водный бассейн. В морях наблюдается также вертикальное зональное распределение температур. Однако эта зональность нарушается течениями.
Охлаждается вода из-за собственного излучения и испарения с поверхности моря. Благодаря постоянному перемешиванию слоев (из-за течений, ветра, конвекционных токов) изменения температур затрагивают значительную толщу воды. Что же касается больших глубин, то там имеется собственный температурный режим.
Благодаря постоянному перемешиванию слоев (из-за течений, ветра, конвекционных токов) изменения температур затрагивают значительную толщу воды. Что же касается больших глубин, то там имеется собственный температурный режим.
Содержание кислорода в морской воде колеблется незначительно. Насыщение им осуществляется в верхних слоях, где живут растения и наблюдается волнение и перемещение вод.
Большое значение имеет соленость морской воды. В открытом океане средняя концентрация растворенных солей равна 3,5 г/л (35 %о), в тропических морях, где сильное испарение, она бывает выше, а в полярных водах ниже, особенно летом (из-за тающих льдов). Соленость морской воды подвержена значительным пространственным и сезонным колебаниям. Существенные изменения ее отражаются на распространении стеногалинных организмов и определяют состав морской фауны. Так, рифообразующие кораллы — типичные стеногалинные формы — крайне чувствительны и к небольшому опреснению воды. Поэтому коралловые рифы прерываются против устьев даже небольших рек. Эврига-линные организмы распространены более широко, чем стеногалинные. В таких морях, как Балтийское, наблюдается закономерное изменение фауны по градиенту солености: от пролива Каттегат до Ботнического залива соленость падает с 32 до 3 %о, и параллельно этому уменьшается число морских видов рыб, моллюсков, раков и др.
Эврига-линные организмы распространены более широко, чем стеногалинные. В таких морях, как Балтийское, наблюдается закономерное изменение фауны по градиенту солености: от пролива Каттегат до Ботнического залива соленость падает с 32 до 3 %о, и параллельно этому уменьшается число морских видов рыб, моллюсков, раков и др.
Важнейшим фактором существования и распространения морских организмов являются течения. Они влияют на распределение температуры в море, смещая его температурные зоны, а также на соленость отдельных участков. Основные океанические течения описывают гигантские круговороты. Различают течения теплые и холодные. Первые возникают в тропической зоне, вторые приносят воду из приполярных областей. Некоторые течения проходят в определенном направлении и постепенно сходят на нет (Гольф-стрим), другие образуют замкнутый круг (Экваториальное проти-вотечение в тропической части Атлантики).
Особое значение течения имеют для пассивно плавающих форм, например мелких ракообразных. Благодаря течениям в центр Атлантики регулярно приносятся и крупные водоросли саргассы с располагающимися на них мшанками, червями, гидроидами, асцидиями и другими животными, обитающими у берегов Южной и Центральной Америки. Крупные морские животные — рыбы, водяные змеи и др. — также регулярно заносятся в новые для них районы океана.
Благодаря течениям в центр Атлантики регулярно приносятся и крупные водоросли саргассы с располагающимися на них мшанками, червями, гидроидами, асцидиями и другими животными, обитающими у берегов Южной и Центральной Америки. Крупные морские животные — рыбы, водяные змеи и др. — также регулярно заносятся в новые для них районы океана.
В море можно выделить две основные группы биотопов в зависимости от того, имеется у них твердый субстрат в виде дна (берега) или нет. Огромную толщу морской воды, где жизнь не связана с твердым субстратом, называют пелагиалью; вся же поверхность дна, включая берег, именуется бенталью (рис. 112).
Обитатели пелагиали в течение всей жизни пассивно или активно плавают. Группа животных, активно передвигающихся благодаря специальным органам, составляет нектон. В нее входят рыбы, китообразные, крупные головоногие моллюски (например, кальмары). Распространены они, как правило, весьма широко, нередко по всему свету. В океанах к таким видам относятся: из млекопитающих касатка, из рыб тунец и некоторые акулы и целый ряд других животных. Организмы, пассивно перемещающиеся, представляют собой планктон. Большинство из них совершает плавательные движения, но они обеспечивают лишь незначительные, в основном вертикальные перемещения, не позволяющие противостоять течениям. Планктонные животные имеют специальные приспособления, уменьшающие массу тела или увеличивающие его поверхность.
Организмы, пассивно перемещающиеся, представляют собой планктон. Большинство из них совершает плавательные движения, но они обеспечивают лишь незначительные, в основном вертикальные перемещения, не позволяющие противостоять течениям. Планктонные животные имеют специальные приспособления, уменьшающие массу тела или увеличивающие его поверхность.
Вертикальное расчленение бентали гораздо сложнее. Здесь различают литоральную и абиссальную зоны. Первая охватывает поверхность континентального шельфа (материковой отмели) от зоны заплеска (участков, не заливающихся водой, но постоянно увлажненных брызгами морской воды) до начала материкового ската. Абиссальная зона включает морское дно и прилегающий к нему слой воды от края материкового ската. Резкой границы между литоралью и абиссалью нет, их разделяет переходная батиальная зона.
Рис. 112. Вертикальное расчленение океана
(по А.В.Константинову, из В.А.Радкевича, 1977)
Область континентального шельфа невелика (лишь 7,5 % площади дна Мирового океана), но чрезвычайно богата животным населением.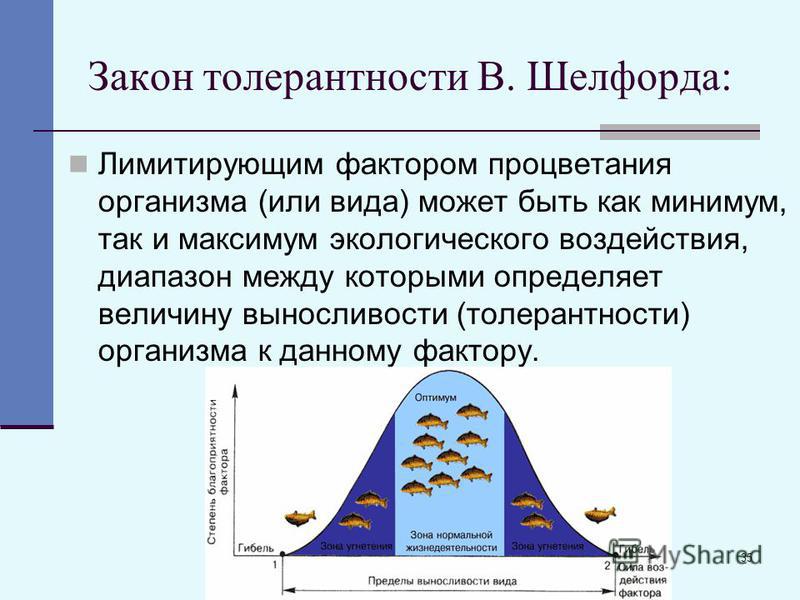 Объясняется это разнообразием биотопов (наличие различных грунтов и зарослей подводных растений, опресненность прилегающих к устьям рек участков и т. д.) и обилием пищи. Пищей служат заросли водорослей и морских трав на мелководьях, где для них имеются хорошие условия освещенности, а также органические вещества, приносимые материковыми водами.
Объясняется это разнообразием биотопов (наличие различных грунтов и зарослей подводных растений, опресненность прилегающих к устьям рек участков и т. д.) и обилием пищи. Пищей служат заросли водорослей и морских трав на мелководьях, где для них имеются хорошие условия освещенности, а также органические вещества, приносимые материковыми водами.
Абиссальная зона — глубоководная часть моря. По площади она во много раз превышает другие зоны, поскольку к ней относится около 80 % морского дна. В отличие от пелагиали здесь постоянно царит тьма, держится ровная температура (в Северной Атлантике 3 «С, в Антарктике 0,5 °С), течения весьма слабые, соленость высокая, грунт очень мягкий, илистый, а давление чрезвычайно велико. Такой комплекс условий почти не меняется на всем протяжении абиссали Мирового океана. Поэтому она считается одной из самых однообразных сред нашей планеты. Пищевые условия абиссали тяжелые. Ресурсы пищи представлены в основном детритом, падающим сверху. Погружаясь, он разлагается бактериями, и до дна доходит лишь небольшая его часть. Вот почему в глубоких водах не бывает изобилия животных. Главными обитателями здесь являются иглокожие. Они питаются илом. Есть на дне и фильтраторы — актинии, черви, моллюски, стеклянные губки, а также крабы и морские пауки-пантоподы, часть из которых де-тритоядные, другие хищники.
Вот почему в глубоких водах не бывает изобилия животных. Главными обитателями здесь являются иглокожие. Они питаются илом. Есть на дне и фильтраторы — актинии, черви, моллюски, стеклянные губки, а также крабы и морские пауки-пантоподы, часть из которых де-тритоядные, другие хищники.
Постоянство условий существования в абиссали привело к тому, что здесь сохранилось много реликтов — древних животных, которые в других участках Мирового океана давно вымерли. Это стебельчатые морские лилии, рыбы-химеры и ряд других. В фауне абиссали есть и представители молодых групп животных, что свидетельствует о заселении абиссали выходцами из пелагиали и ли-торали.
Экологическая валентность, или экологическая пластичность, или экологическая толерантность
Пользователи также искали:
экологическая пластичность,
экологическая толерантность,
экологическая валентность это,
экологическая валентность примеры,
экологическая валентность вида по отношению к экологической валентности отдельной особи,
экологический спектр вида,
какова экологическая валентность водных обитателей,
понятие экологической валентности,
Экологическая,
экологическая,
валентность,
Экологическая валентность,
валентности,
экологической,
вида,
понятие экологической валентности,
экологический спектр вида,
экологическая пластичность,
экологическая толерантность,
примеры,
какова,
водных,
обитателей,
отношению,
отдельной,
особи,
понятие,
экологический,
спектр,
пластичность,
толерантность,
экологическая валентность это,
экологическая валентность примеры,
какова экологическая валентность водных обитателей,
экологическая валентность вида по отношению к экологической валентности отдельной особи,
экологическая валентность,
экологические термины. экологическая валентность,
экологическая валентность,
…
Indicator signs of the adaptation of subtropical wood plants based on complex researches
Для Нижнего Поволжья со сложными почвенноклиматическими условиями прогнозы несут рост продолжительности вегетации, увеличение тепла, смягчение зим. Это указывает на необходимость исследований субтропических растений для объективной оценки выявления механизмов их адаптации и перспективности для создания защитных лесных насаждений в малолесных регионах. Теоретический и практический интерес для Волгоградской области представляет ценное плодовое, лекарственное, лесомелиоративное и декоративное растениекустарник (унаби) Zizyphus jujubа Mill. (семейство Rhamnaceae). Цель исследований изучить экологофизиологические особенности субтропических древесных растений Zizyphus jujubа и выявить индикаторные признаки их адаптации в Волгоградской области. Объектами исследований являлись сортовые растения унаби (Таянцзао, Южанин крупноплодные, Дружба, Финик среднеплодные, Сочинский, Темрюкский мелкоплодные). Они испытываются в Волгоградской области впервые, получены из ФГБНУ ВНИИЦиСК (г. Сочи) и возделываются в ФГБНУ ФНЦ агроэкологии РАН (Волгоградская область, Россия). Почвы экспериментального участка (504505с.ш. и 45224523 в.д.) характеризуются небольшим содержанием гумуса (с 0,57 до 1,15 ). Выявлено, что для района исследований (Волгоградская область) характерны небольшое количество осадков, низкие температуры зимой, высокие летом, засухи высокой и средней интенсивности. Амплитуда абсолютных максимумов и минимумов варьирует от семидесяти восьми (сухая степь) до девяноста градусов по Цельсию (полупустыня). Установлено, что агроклиматические ресурсы ареалов естественного распространения отличаются от пунктов введения Zizyphus jujubа Mill. в культуру. Разные уровни морфологической изменчивости, а также водообеспеченности приводят к изменению способности расходовать и удерживать воду, к дефициту влаги в листьях у Z.jujubа. Незначительное влияние засухи на состояние надземных органов растений в острозасушливые периоды отмечено у среднеплодных и крупноплодных сортов.
Они испытываются в Волгоградской области впервые, получены из ФГБНУ ВНИИЦиСК (г. Сочи) и возделываются в ФГБНУ ФНЦ агроэкологии РАН (Волгоградская область, Россия). Почвы экспериментального участка (504505с.ш. и 45224523 в.д.) характеризуются небольшим содержанием гумуса (с 0,57 до 1,15 ). Выявлено, что для района исследований (Волгоградская область) характерны небольшое количество осадков, низкие температуры зимой, высокие летом, засухи высокой и средней интенсивности. Амплитуда абсолютных максимумов и минимумов варьирует от семидесяти восьми (сухая степь) до девяноста градусов по Цельсию (полупустыня). Установлено, что агроклиматические ресурсы ареалов естественного распространения отличаются от пунктов введения Zizyphus jujubа Mill. в культуру. Разные уровни морфологической изменчивости, а также водообеспеченности приводят к изменению способности расходовать и удерживать воду, к дефициту влаги в листьях у Z.jujubа. Незначительное влияние засухи на состояние надземных органов растений в острозасушливые периоды отмечено у среднеплодных и крупноплодных сортов. Повреждаются молодые неодревесневшие побеги (при температуре воздуха около 40 С и снижении влажности воздуха до 10). Снижение тургора листьев наблюдалось при показателях водного дефицита 28,335,8 . Водный дефицит снижается на 610 с увеличением возраста, что обусловлено повышением адаптации и регуляции водного баланса. По показателям относительного выхода электролитов определены группы по степени засухоустойчивости: низкая Южанин, ТаЯнЦзао (4,234,71), средняя Финик, Дружба (3,103,61) и высокая Темрюкский, Сочинский (1,641,99). Экологическая толерантность различных организмов к низким температурам достаточно специфична. Оценка различных сортов Zizyphus jujubа в условиях светлокаштановых почв показала, что адаптация растений к низким температурам повышается с увеличением возраста. У мелкоплодных сортов с увеличением возраста повреждения незначительны. По материалам вегетационного опыта установлены пределы экологической толерантности мелкоплодных форм Zizyphus jujubа к хлоридному засолению, что дает возможность прогнозировать их успешное выращивание на почвах с содержанием ионов хлора 0,1.
Повреждаются молодые неодревесневшие побеги (при температуре воздуха около 40 С и снижении влажности воздуха до 10). Снижение тургора листьев наблюдалось при показателях водного дефицита 28,335,8 . Водный дефицит снижается на 610 с увеличением возраста, что обусловлено повышением адаптации и регуляции водного баланса. По показателям относительного выхода электролитов определены группы по степени засухоустойчивости: низкая Южанин, ТаЯнЦзао (4,234,71), средняя Финик, Дружба (3,103,61) и высокая Темрюкский, Сочинский (1,641,99). Экологическая толерантность различных организмов к низким температурам достаточно специфична. Оценка различных сортов Zizyphus jujubа в условиях светлокаштановых почв показала, что адаптация растений к низким температурам повышается с увеличением возраста. У мелкоплодных сортов с увеличением возраста повреждения незначительны. По материалам вегетационного опыта установлены пределы экологической толерантности мелкоплодных форм Zizyphus jujubа к хлоридному засолению, что дает возможность прогнозировать их успешное выращивание на почвах с содержанием ионов хлора 0,1. Таким образом, проведенные комплексные экологофизиологические исследования адаптационной способности субтропических растений Zizyphus jujubа выявили индикаторные признаки, позволяющие оценивать и прогнозировать степень адаптации в зависимости от сортовой принадлежности и реакцию на стрессовые факторы, специфичные для условий Нижнего Поволжья.
Таким образом, проведенные комплексные экологофизиологические исследования адаптационной способности субтропических растений Zizyphus jujubа выявили индикаторные признаки, позволяющие оценивать и прогнозировать степень адаптации в зависимости от сортовой принадлежности и реакцию на стрессовые факторы, специфичные для условий Нижнего Поволжья.
For the Lower Volga region with difficult soil and climatic conditions, forecasts bear an increase in the length of the growing season, an increase in heat, and a softer winters. This indicates the need for studies of subtropical plants for an objective assessment of the identification of mechanisms for their adaptation and prospects for creating protective forest plantations in sparsely wooded regions. Theoretical and practical interest for the Volgograd region is a valuable fruit, medicinal, forest reclamation and ornamental shrub (unabi) plant Zizyphus jujuba Mill. (family Rhamnaceae). The purpose of the research is to study the ecological and physiological features of subtropical woody plants of Zizyphus jujuba and to identify indicator signs of their adaptation in the Volgograd region. The objects of research were the varietal plants of the unabi (Tayangzao, Southerner largefruited, Druzhba, Phenicia mediumfruited, Sochi, Temryuk smallfruited). They are tested in the Volgograd region for the first time, obtained from the FSBI (city of Sochi) and are cultivated in FSBI FNTS Agroecology RAS (Volgograd region, Russia). The soils of the experimental plot (50 450 5N and 45 2245 23E) are characterized by a low humus content (from 0.57 to 1.15). It was revealed that the study area (Volgograd region) is characterized by a small amount of precipitation, low temperatures in winter, high temperatures in summer, droughts of high and medium intensity. The amplitude of the absolute maxima and minima varies from seventyeight (dry steppe) to ninety degrees Celsius (semidesert). It has been established that the agroclimatic resources of natural distribution areas differ from the introduction points of Zizyphus jujuba Mill. to culture. Different levels of morphological variability, as well as water availability, lead to a change in the ability to expend and retain water, to a lack of moisture in the leaves of Z.
The objects of research were the varietal plants of the unabi (Tayangzao, Southerner largefruited, Druzhba, Phenicia mediumfruited, Sochi, Temryuk smallfruited). They are tested in the Volgograd region for the first time, obtained from the FSBI (city of Sochi) and are cultivated in FSBI FNTS Agroecology RAS (Volgograd region, Russia). The soils of the experimental plot (50 450 5N and 45 2245 23E) are characterized by a low humus content (from 0.57 to 1.15). It was revealed that the study area (Volgograd region) is characterized by a small amount of precipitation, low temperatures in winter, high temperatures in summer, droughts of high and medium intensity. The amplitude of the absolute maxima and minima varies from seventyeight (dry steppe) to ninety degrees Celsius (semidesert). It has been established that the agroclimatic resources of natural distribution areas differ from the introduction points of Zizyphus jujuba Mill. to culture. Different levels of morphological variability, as well as water availability, lead to a change in the ability to expend and retain water, to a lack of moisture in the leaves of Z. jujuba. The insignificant effect of drought on the state of the aboveground plant organs during the arid periods was observed in mediumsized and largefruited varieties. Young, nonlignified shoots are damaged (at an air temperature of about 40 C and a decrease in air humidity of up to 10). A decrease in leaf turgor was observed with water deficit indicators of 28.335.8. Water deficiency decreases by 610 with increasing age, which is due to increased adaptation and regulation of water balance. According to the indicators of the relative electrolyte yield, groups were determined according to the degree of drought resistance: low Southerner, TaYanZao (4.234.71), medium Finik, Friendship (3.103.61) and high Temryuk, Sochi (1.641.99). Ecological tolerance of various organisms to low temperatures is quite specific. Evaluation of different varieties of Zizyphus jujuba under lightbrown soils showed that the adaptation of plants to low temperatures increases with increasing age. In small varieties with an increase in age, the damage is minor.
jujuba. The insignificant effect of drought on the state of the aboveground plant organs during the arid periods was observed in mediumsized and largefruited varieties. Young, nonlignified shoots are damaged (at an air temperature of about 40 C and a decrease in air humidity of up to 10). A decrease in leaf turgor was observed with water deficit indicators of 28.335.8. Water deficiency decreases by 610 with increasing age, which is due to increased adaptation and regulation of water balance. According to the indicators of the relative electrolyte yield, groups were determined according to the degree of drought resistance: low Southerner, TaYanZao (4.234.71), medium Finik, Friendship (3.103.61) and high Temryuk, Sochi (1.641.99). Ecological tolerance of various organisms to low temperatures is quite specific. Evaluation of different varieties of Zizyphus jujuba under lightbrown soils showed that the adaptation of plants to low temperatures increases with increasing age. In small varieties with an increase in age, the damage is minor. Based on the vegetation experience, the limits of ecological tolerance of smallfruited forms of Zizyphus jujuba to chloride salinity were established, which makes it possible to predict their successful cultivation on soils with a chlorine ion content of 0.1. Thus, complex ecological and physiological studies of the adaptive capacity of subtropical plants Zizyphus jujuba revealed indicator signs that allow to evaluate and predict the degree of adaptation depending on the variety and the response to stress factors specific to the conditions of the Lower Volga region.
Based on the vegetation experience, the limits of ecological tolerance of smallfruited forms of Zizyphus jujuba to chloride salinity were established, which makes it possible to predict their successful cultivation on soils with a chlorine ion content of 0.1. Thus, complex ecological and physiological studies of the adaptive capacity of subtropical plants Zizyphus jujuba revealed indicator signs that allow to evaluate and predict the degree of adaptation depending on the variety and the response to stress factors specific to the conditions of the Lower Volga region.
Экологический допуск
по JSTOR
Теория выражения реакции населения на независимые от плотности градиенты факторов окружающей среды выведена для случая асексуальности. Показано, что экологическая толерантность генотипа является функцией по крайней мере четырех параметров: g $ _1 $ и V $ _ {E1} $, экологического оптимума и его вариации в развитии между людьми, а также g $ _2 $ и V $. _ {E2} $, ожидаемый генетический вклад в широту адаптации и ее вариативность в развитии. {1/2} $ может развиваться по-разному g $ _2 $, если существуют явные межвидовые различия для V $ _ {E1} $ и V $ _ {E2} $. Показано, что метод максимального правдоподобия способен генерировать точные оценки генотипических параметров g $ _1 $, g $ _2 $, V $ _ {E1} $ и V $ _ {E2} $ с умеренно большими выборками. Мы предлагаем, как эту процедуру можно использовать для оценки аналогичных параметров для популяции смешанных генотипов и для получения оценок генетической изменчивости для оптимума окружающей среды и широты адаптации.Указывается на потенциальную полезность этой методологии для анализа данных, обычно генерируемых в программах экологической оценки и селекции растений.
{1/2} $ может развиваться по-разному g $ _2 $, если существуют явные межвидовые различия для V $ _ {E1} $ и V $ _ {E2} $. Показано, что метод максимального правдоподобия способен генерировать точные оценки генотипических параметров g $ _1 $, g $ _2 $, V $ _ {E1} $ и V $ _ {E2} $ с умеренно большими выборками. Мы предлагаем, как эту процедуру можно использовать для оценки аналогичных параметров для популяции смешанных генотипов и для получения оценок генетической изменчивости для оптимума окружающей среды и широты адаптации.Указывается на потенциальную полезность этой методологии для анализа данных, обычно генерируемых в программах экологической оценки и селекции растений.
Текущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. С момента своего создания в 1867 году журнал The American Naturalist сохраняет свои позиции в качестве одной из ведущих рецензируемых публикаций в мире по вопросам экологии, эволюции и исследований поведения. Его цели — публиковать статьи, представляющие широкий интерес для читателей, ставить новые и важные проблемы, знакомить с новыми предметами, развивать концептуальную унификацию и изменять образ мышления людей.AmNat делает упор на сложные методологии и новаторские теоретические синтезы, стремясь расширить знания об органической эволюции и других общих биологических принципах.
Его цели — публиковать статьи, представляющие широкий интерес для читателей, ставить новые и важные проблемы, знакомить с новыми предметами, развивать концептуальную унификацию и изменять образ мышления людей.AmNat делает упор на сложные методологии и новаторские теоретические синтезы, стремясь расширить знания об органической эволюции и других общих биологических принципах.
С момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета, University of Chicago Press взяла на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, которые способствуют образованию и способствуют пониманию общества. , и обогатить культурную жизнь.Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также естественные науки.
Перейти к основному содержанию
Поиск
Поиск
- Где угодно
- Быстрый поиск где угодно
Поиск Поиск
Расширенный поиск
Войти | регистр
Пропустить основную навигацию Закрыть меню ящика Открыть меню ящика Домой
- Подписка / продление
- Учреждения
- Индивидуальные подписки
- Индивидуальные продления
- Библиотекари
- Тарифы, заказы и платежи
- Завершено Чикагский пакет
- Полный цикл и охват содержимого
- Файлы KBART и RSS-каналы
- Разрешения и перепечатки
- Инициатива развивающихся стран Чикаго
- Даты отправки и заявки
- Часто задаваемые вопросы библиотекарей
- Агенты
- Тарифы, заказы, и платежи
- Полный пакет Chicago
- Полный охват и содержание
- Даты отправки и претензии
- Часто задаваемые вопросы об агенте
- Партнеры по издательству
- О нас
- Публикуйте с нами
- Недавно приобретенные журналы
- Издательское право tners
- Новости прессы
- Подпишитесь на оповещения eTOC
- Пресс-релизы
- Медиа
- Книги издательства Чикагского университета
- Распределительный центр в Чикаго
- Чикагский университет
- Положения и условия
- Заявление о публикационной этике
- Уведомление о конфиденциальности
- Доступность Chicago Journals
- Доступность университета
- Следуйте за нами на facebook
- Следуйте за нами в Twitter
- Свяжитесь с нами
- Медиа и рекламные запросы
- Открытый доступ в Чикаго
- Следуйте за нами на facebook
- Следуйте за нами в Twitter
Ниша
Ниша
Ниша
Для любого фактора окружающей среды (например,грамм. DO, pH, сила света, ток
DO, pH, сила света, ток
скорость и т. д.) будет диапазон, в котором конкретный вид
могу терпеть.
Организм с узким диапазоном толерантности считается «стеноз ..»
для этого фактора (например, стенотермический или стеногалинный). Организм с
считается, что широкий диапазон допуска составляет «эври ..» (например, эвритермический).
Организмы могут быть стенозированы по одному фактору и эври по отношению
к другому. Центрархиды (окунь и солнечная рыба) эвритермны, но
стеногалин; лососевые — как раз наоборот.Эти диапазоны допусков не являются независимыми. Организм около толерантности
ограничения для одного фактора, вероятно, будут испытывать стресс, поэтому его способность
переносимость других факторов будет снижена. Диапазоны допусков для окружающей среды
факторы частично определяют нишу организма. Если устойчивость к солености нанесена на график
против температурной устойчивости определяется площадь ниши.
Отображение третьего фактора (например, pH) под прямым углом к первым двум создает
объем ниши.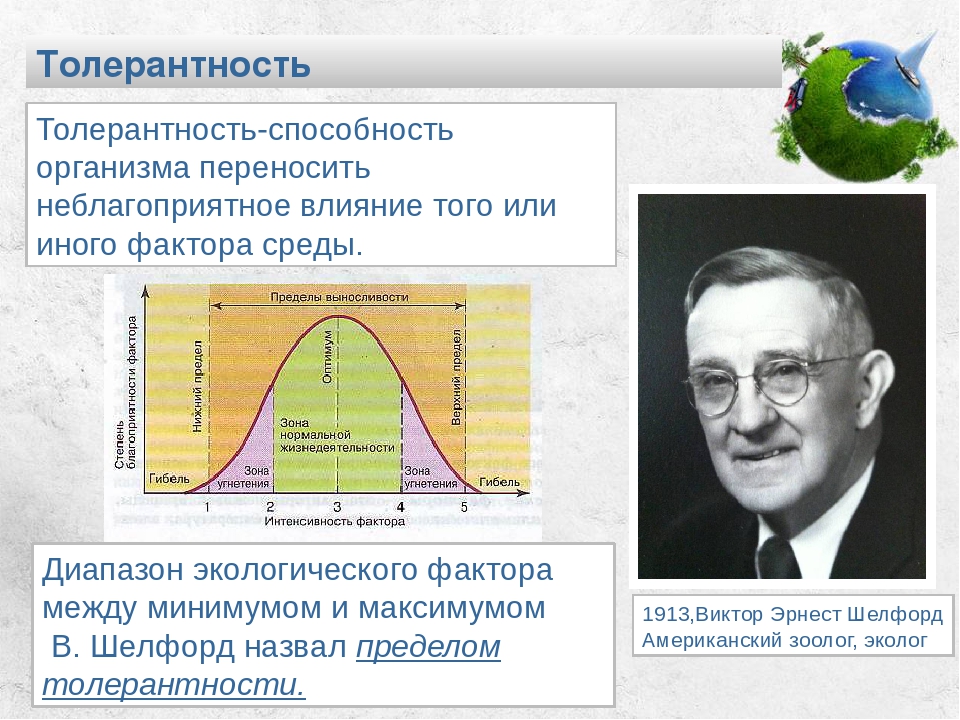 Если каждый из дополнительных диапазонов допуска, включая не
Если каждый из дополнительных диапазонов допуска, включая не
только физические факторы, но также и биотические факторы, такие как тип пищи и
восприимчивости хищников, отображается под прямым углом ко всем остальным,
результатом является (теоретически) n-мерный гиперобъем , как предлагается
автор Г.Э. Хатчинсон как определение ниши .
Допустимый диапазон, показываемый видом для определенного фактора, не является
абсолютное, как мы видели из взаимосвязи между факторами. В
на допустимый диапазон также влияет недавняя история болезни организма;
он может переносить более высокие температуры, если привык к ним (адаптироваться к ним).
Пространство, определяемое всеми диапазонами допусков для разной акклиматизации
температура известна как многоугольник допуска температуры.Толерантность
полигоны характерны для рассматриваемых видов и могут быть
сильно отличается для других видов.
относительный. Наступает точка (максимальная или минимальная), за которой
организм не может акклиматизироваться. Организмы могут адаптироваться к окружающей среде.
Организмы могут адаптироваться к окружающей среде.
факторы, кроме температуры, даже включая токсичные вещества. Предположительно
для любого фактора окружающей среды существует полигон допусков.
Однако допуски не остаются постоянными на протяжении всего жизненного цикла
организм.Полигоны толерантности для яиц или молоди могут быть совершенно разными.
чем для взрослых. Для большинства организмов условия, при которых они могут
размножение намного более ограничено, чем условия, при которых взрослые могут
выживать. Разумно визуализировать репродуктивный многоугольник внутри
полигон общего допуска.
Адаптировано со страницы в Niche Space.
Нажмите кнопку «Назад», чтобы вернуться к уроку.
Границы | Информация о цветущих медузах (класс: Scyphozoa) в Мексиканском заливе: диапазоны и пределы экологической толерантности указывают на различия в предпочтениях сред обитания и устойчивости к изменению климата среди их представителей
Введение
Медузы — важные компоненты морских пищевых сетей. Они питаются зоопланктоном и личинками рыб (Möller, 1984; Purcell and Sturdevant, 2000; Riascos et al., 2014) и служат пищей для различных морских животных, таких как пингвины, черепахи и тунец (Hays et al., 2018). . Крупные скопления медуз, также известные как «цветение», связаны с многочисленными негативными социально-экономическими последствиями. Медузы забивают рыболовные сети (Nagata et al., 2009), снижают качество улова (Quiñones et al., 2013), затрудняют водозаборники для охлаждения электростанций (Abdul Azis et al., 2000) и кусают посетителей пляжа (De Donno et al., 2014). Цветение также создает проблемы для аквакультуры, засоряя загоны сети, а слизь, богатая нематоцистами медуз, ответственна за нарушения жабр рыб (Purcell et al., 2013).
Они питаются зоопланктоном и личинками рыб (Möller, 1984; Purcell and Sturdevant, 2000; Riascos et al., 2014) и служат пищей для различных морских животных, таких как пингвины, черепахи и тунец (Hays et al., 2018). . Крупные скопления медуз, также известные как «цветение», связаны с многочисленными негативными социально-экономическими последствиями. Медузы забивают рыболовные сети (Nagata et al., 2009), снижают качество улова (Quiñones et al., 2013), затрудняют водозаборники для охлаждения электростанций (Abdul Azis et al., 2000) и кусают посетителей пляжа (De Donno et al., 2014). Цветение также создает проблемы для аквакультуры, засоряя загоны сети, а слизь, богатая нематоцистами медуз, ответственна за нарушения жабр рыб (Purcell et al., 2013).
В Мексиканском заливе (GoM) проблемное цветение медуз в прибрежных районах часто вызывается медузами класса Scyphozoa, Phylum Cnidaria. Сцифозойные медузы имеют мультимодальный жизненный цикл (Ceh et al., 2015), при котором многолетний бентосный полип производит сезонных медуз.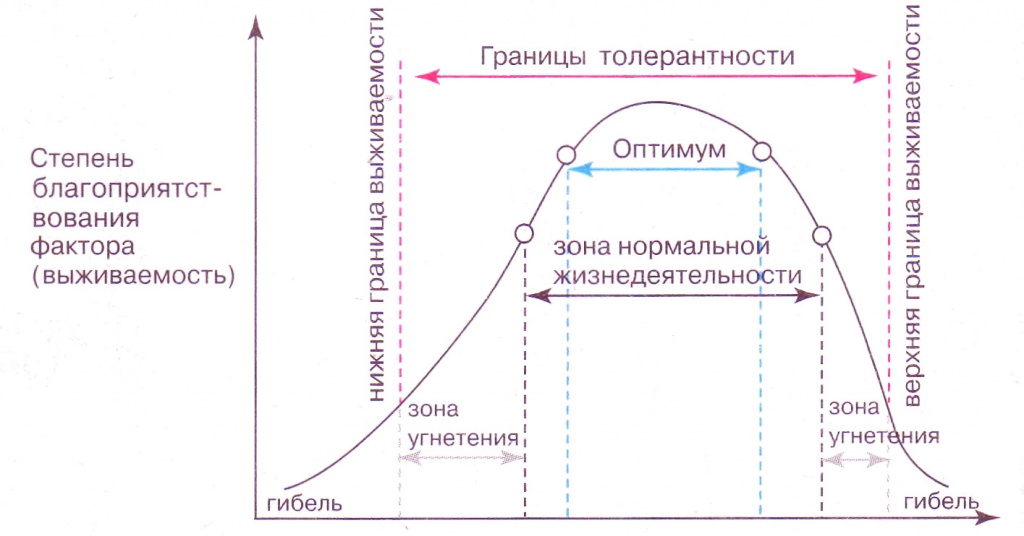 Молодые медузы (эфиры) продуцируются полипами в процессе бесполого поперечного деления, называемого «стробиляцией».В Правительстве Молдовы зарегистрировано 20 видов Scyphozoa, представляющих три отряда и одиннадцать семейств (Segura-Puertas et al., 2009). Преобладающими родами цветущих медуз в прибрежных и шельфовых экосистемах являются Aurelia , Chrysaora и Stomolophus (Larson, 1991), которые цветут в основном в летние месяцы (Graham, 2001; Robinson and Graham, 2013).
Молодые медузы (эфиры) продуцируются полипами в процессе бесполого поперечного деления, называемого «стробиляцией».В Правительстве Молдовы зарегистрировано 20 видов Scyphozoa, представляющих три отряда и одиннадцать семейств (Segura-Puertas et al., 2009). Преобладающими родами цветущих медуз в прибрежных и шельфовых экосистемах являются Aurelia , Chrysaora и Stomolophus (Larson, 1991), которые цветут в основном в летние месяцы (Graham, 2001; Robinson and Graham, 2013).
Полипы играют ключевую роль в поддержании и расширении популяций скифозов (Lucas et al., 2012).Каждый полип высвобождает несколько медуз за одно событие стробиляции. Производство медуз контролируется количеством стробилирующих полипов, а также скоростью и продолжительностью выпуска медуз (Lucas et al., 2012), поэтому размер цветения медуз частично определяется размером популяции полипов. Полипы также размножаются бесполым путем, увеличивая размер бентосной популяции и тем самым способствуя масштабам цветения медуз (Lucas et al. , 2012).
, 2012).
Текущие знания о расположении, размере и динамике естественных полипов в GoM отсутствуют.Полипы крошечные и находятся в защищенных, плохо видимых местах, что затрудняет их обнаружение. Известно, что полипы обитают на твердых субстратах, включая биообрастающие бентосные организмы, плавучие платформы и искусственные сооружения (Duarte et al., 2013). Дно большей части МП имеет мягкое песчаное или илистое дно, поэтому площади поселений, вероятно, ограничены. Однако, несмотря на заметное цветение, полипы даже самых распространенных сцифозных видов не были обнаружены в GoM. Невозможность локализации популяций полипов в природе затрудняет изучение динамики и цветения популяций медуз.Например, триггеры стробиляции и образования медуз не могут быть изучены in situ , а географическое происхождение цветения медуз неизвестно. Более того, без знания нынешних географических ареалов популяций полипов трудно предсказать, как медузы отреагируют на изменение климата.
Реакция скифозов на изменение климата до конца не изучена. Присутствие и численность медуз в экосистемах, как правило, сильно варьируется. Время, местоположение и количество наблюдаемых особей могут значительно различаться в зависимости от года и местоположения (Purcell, 2005; Heim-Ballew and Olsen, 2019).Изменчивость частоты и масштабов цветения медуз частично объясняется глобальными многомесячными климатическими колебаниями (Condon et al., 2014). Однако данные некоторых экосистем позволяют предположить, что антропогенные нарушения экосистем могут способствовать формированию цветения (Purcell et al., 2007; Purcell, 2011). Было показано, что численность медузы увеличивается в экосистемах, где интенсивно ведется рыбный промысел (Lynam et al., 2006) в районах с бентосной гипоксией (Shoji et al., 2010; Miller and Graham, 2012) и в районах, подверженных эвтрофикации (Purcell et al., 1999a; Haraldsson et al., 2012).
Присутствие и численность медуз в экосистемах, как правило, сильно варьируется. Время, местоположение и количество наблюдаемых особей могут значительно различаться в зависимости от года и местоположения (Purcell, 2005; Heim-Ballew and Olsen, 2019).Изменчивость частоты и масштабов цветения медуз частично объясняется глобальными многомесячными климатическими колебаниями (Condon et al., 2014). Однако данные некоторых экосистем позволяют предположить, что антропогенные нарушения экосистем могут способствовать формированию цветения (Purcell et al., 2007; Purcell, 2011). Было показано, что численность медузы увеличивается в экосистемах, где интенсивно ведется рыбный промысел (Lynam et al., 2006) в районах с бентосной гипоксией (Shoji et al., 2010; Miller and Graham, 2012) и в районах, подверженных эвтрофикации (Purcell et al., 1999a; Haraldsson et al., 2012).
Ожидается, что в Правительстве страны изменение климата повлияет на температуру, характер осадков, активность тропических штормов и уровень моря (Biasutti et al. , 2012). Повышение температуры дна может повлиять на бентические полипы, их выживаемость и скорость их стробиляции, что повлияет на величину и частоту цветения. Температура придонной воды повысилась на 2 ° C за 30-летний период на северном континентальном шельфе правительства штата Мичиган (Turner et al., 2017), что составляет 1.В 9 раз быстрее, чем местное повышение температуры воздуха в летние месяцы, и в 6,4 раза быстрее, чем глобальное годовое повышение температуры моря (Turner et al., 2017). Кроме того, средняя температура воды GoM, по прогнозам, увеличится на 4 ° C к концу века (Muhling et al., 2011; Biasutti et al., 2012). Как это повышение температуры повлияет на популяции скифозов и частоту цветения, неясно. Было показано, что температура влияет на скорость роста, бесполое размножение и стробиляцию полипов Aurelia spp.(Purcell, 2007; Willcox et al., 2007; Hubot et al., 2017). Пределы термостойкости ограничивают биогеографический диапазон, в котором могут выжить скифозные виды.
, 2012). Повышение температуры дна может повлиять на бентические полипы, их выживаемость и скорость их стробиляции, что повлияет на величину и частоту цветения. Температура придонной воды повысилась на 2 ° C за 30-летний период на северном континентальном шельфе правительства штата Мичиган (Turner et al., 2017), что составляет 1.В 9 раз быстрее, чем местное повышение температуры воздуха в летние месяцы, и в 6,4 раза быстрее, чем глобальное годовое повышение температуры моря (Turner et al., 2017). Кроме того, средняя температура воды GoM, по прогнозам, увеличится на 4 ° C к концу века (Muhling et al., 2011; Biasutti et al., 2012). Как это повышение температуры повлияет на популяции скифозов и частоту цветения, неясно. Было показано, что температура влияет на скорость роста, бесполое размножение и стробиляцию полипов Aurelia spp.(Purcell, 2007; Willcox et al., 2007; Hubot et al., 2017). Пределы термостойкости ограничивают биогеографический диапазон, в котором могут выжить скифозные виды. Способность переносить региональные или местные термические условия также может повлиять на вероятность того, что вид станет экзотическим захватчиком. Например, инвазивный Scyphozoan Aurelia coerulea до сих пор укоренился в местообитаниях, которые имеют сезонные максимумы и минимумы, аналогичные его естественному диапазону широт от 30 ° до 45 ° северной широты (Dawson et al., 2005; Скоррано и др., 2017).
Способность переносить региональные или местные термические условия также может повлиять на вероятность того, что вид станет экзотическим захватчиком. Например, инвазивный Scyphozoan Aurelia coerulea до сих пор укоренился в местообитаниях, которые имеют сезонные максимумы и минимумы, аналогичные его естественному диапазону широт от 30 ° до 45 ° северной широты (Dawson et al., 2005; Скоррано и др., 2017).
Соленость — еще один важный фактор окружающей среды, который может влиять на развитие и выживание полипов скифозойных медуз (Rippingale and Kelly, 1995; Purcell et al., 1999b, 2009; Pitt and Kingsford, 2003). Вспышки медуз часто происходят в прибрежных средах с переменной соленостью, таких как заливы, эстуарии и частично замкнутые морские воды во всем мире (Purcell et al., 1999b). Кроме того, согласно прогнозам, изменение количества осадков приведет к изменению солености прибрежных районов, в том числе Правительства Молдовы (Biasutti et al., 2012), мотивируя исследования предпочтений солености и ограничений видов медуз. Лишь несколько естественных местообитаний полипов Aurelia spp. были изучены (Gröndahl, 1988; Purcell et al., 2009; Malej et al., 2012; Marques et al., 2015, 2019; Hoèvar et al., 2018), таким образом, появилась информация о разнообразии толерантности к солености в пределах рода ограничено. Многие прошлые исследования также были омрачены наличием множества загадочных видов в пределах рода Aurelia (Dawson and Martin, 2001; Scorrano et al., 2017). Полевые и лабораторные исследования показывают, что конгенеров Aurelia по-разному реагируют на изменение солености (Spangenberg, 1964; Purcell et al., 2009; Marques et al., 2019). На размер диких популяций полипов A. coerulea оказала негативное влияние высокая соленость, особенно в сочетании с высокими температурами (Marques et al., 2019). Низкая соленость задерживала рост диких особей A. labiata (Purcell et al., 2009), в то время как изменение солености в пределах диапазона местных колебаний окружающей среды не оказывало существенного влияния на рост полипов у Aurelia sp.
Лишь несколько естественных местообитаний полипов Aurelia spp. были изучены (Gröndahl, 1988; Purcell et al., 2009; Malej et al., 2012; Marques et al., 2015, 2019; Hoèvar et al., 2018), таким образом, появилась информация о разнообразии толерантности к солености в пределах рода ограничено. Многие прошлые исследования также были омрачены наличием множества загадочных видов в пределах рода Aurelia (Dawson and Martin, 2001; Scorrano et al., 2017). Полевые и лабораторные исследования показывают, что конгенеров Aurelia по-разному реагируют на изменение солености (Spangenberg, 1964; Purcell et al., 2009; Marques et al., 2019). На размер диких популяций полипов A. coerulea оказала негативное влияние высокая соленость, особенно в сочетании с высокими температурами (Marques et al., 2019). Низкая соленость задерживала рост диких особей A. labiata (Purcell et al., 2009), в то время как изменение солености в пределах диапазона местных колебаний окружающей среды не оказывало существенного влияния на рост полипов у Aurelia sp. из Тасмании (Willcox et al., 2007).
из Тасмании (Willcox et al., 2007).
Диапазоны, пределы допустимости температуры и солености и способность к акклиматизации сильно влияют на распространение морских видов (Pörtner, 2002; Stillman, 2003; Somero, 2005, 2010), но неизвестны для большинства видов медуз. Тем не менее, пределы толерантности имеют решающее значение для понимания современного распределения полипов медуз в ПМ и того, как это распределение может измениться в сценариях изменения климата. В этом исследовании с помощью лабораторных экспериментов мы оценили устойчивость к температуре и солености полипов двух видов: Aurelia , собранных в Правительстве США, и инвазивного вида Aurelia , обитающего в Южном и Восточно-Китайском морях (Dawson et al., 2005). А именно, мы ориентируемся на Aurelia sp. 9 и новый вид Aurelia , о котором впервые сообщается в этой статье и который был обнаружен в прибрежных водах правительства штата Мичиган, а также Aurelia coerulea , вид, родом из Японии, который вторгся на тихоокеанское побережье Соединенных Штатов и другие районы вокруг мире (Dawson et al. , 2005; Scorrano et al., 2017). Наши цели: 1) определить диапазон и пределы температур, которые каждый вид может выдерживать в природе, 2) выяснить, имеют ли три вида одинаковые или разные верхние тепловые пределы и 3) определить диапазоны и пределы допустимости солености для каждого вида .Это исследование направлено на определение пределов толерантности к температуре и солености для трех видов Aurelia , прогноз их биогеографического распределения в Правительстве штата Мичиган и обеспечение понимания того, как популяции медуз могут жить при повышении температуры океана.
, 2005; Scorrano et al., 2017). Наши цели: 1) определить диапазон и пределы температур, которые каждый вид может выдерживать в природе, 2) выяснить, имеют ли три вида одинаковые или разные верхние тепловые пределы и 3) определить диапазоны и пределы допустимости солености для каждого вида .Это исследование направлено на определение пределов толерантности к температуре и солености для трех видов Aurelia , прогноз их биогеографического распределения в Правительстве штата Мичиган и обеспечение понимания того, как популяции медуз могут жить при повышении температуры океана.
Материалы и методы
Учреждение источников организмов и культур
Пять самок медуз Aurelia sp. 9 были собраны в заливе Галвестон в октябре 2017 года. Медузы несли планулы.Вскоре после сбора образцов планулы были изолированы и помещены в контейнеры емкостью 700 мл с фильтрованной морской водой солености окружающей среды. Планулы были доставлены в Техасский университет A&M в Галвестонский центр морской жизни, где они были объединены в единую культуру и позволили трансформироваться в полипы. Приблизительно 50 полипов, принадлежащих к виду A. coerulea , были предоставлены аквариумом Moody Gardens и использовались для создания культур. Одна живая взрослая самка медузы Aurelia , несущая планулы, была обнаружена сачком 1 июля 2017 года во время исследовательского рейса на борту НИС Pelican .Сбор произошел примерно в 80 милях к югу от побережья Луизианы в провинции Мичиган (28 ° 0 ′ 0 ″ северной широты, –89 ° 4 ′ 8 ″ з.д.). Приборы на борту исследовательского судна измерили параметры воды: соленость 37 ppt и температура 28,8 ° C. Ткань медузы сохранялась в 100% этаноле. Планулы были собраны с медузы и доставлены в Техасский университет A&M в Галвестоне, где они превратились в полипы. Полипы всех трех видов содержались в Sea Life Facility Техасского университета A&M в Галвестоне в вентилируемых аквариумах при солености 33–35 ppt, температуре окружающей среды 15–23 ° C, минимальном освещении и кормление один или два раза в неделю комбинация только что вылупившихся науплий Artemia salina и обогащенных водорослями коловраток.
Приблизительно 50 полипов, принадлежащих к виду A. coerulea , были предоставлены аквариумом Moody Gardens и использовались для создания культур. Одна живая взрослая самка медузы Aurelia , несущая планулы, была обнаружена сачком 1 июля 2017 года во время исследовательского рейса на борту НИС Pelican .Сбор произошел примерно в 80 милях к югу от побережья Луизианы в провинции Мичиган (28 ° 0 ′ 0 ″ северной широты, –89 ° 4 ′ 8 ″ з.д.). Приборы на борту исследовательского судна измерили параметры воды: соленость 37 ppt и температура 28,8 ° C. Ткань медузы сохранялась в 100% этаноле. Планулы были собраны с медузы и доставлены в Техасский университет A&M в Галвестоне, где они превратились в полипы. Полипы всех трех видов содержались в Sea Life Facility Техасского университета A&M в Галвестоне в вентилируемых аквариумах при солености 33–35 ppt, температуре окружающей среды 15–23 ° C, минимальном освещении и кормление один или два раза в неделю комбинация только что вылупившихся науплий Artemia salina и обогащенных водорослями коловраток. Морская вода соответствующей солености была получена путем добавления аквариумной соли Instant Ocean к фильтрованной морской воде с соленостью окружающей среды залива до достижения целевой солености. Воду в аквариумах меняли раз в неделю. Вторая частичная медуза Aurelia была собрана 3 июля 2017 года в нейстонную сеть и сохранена в 100% этаноле. Оба образца медуз были использованы для молекулярного анализа.
Морская вода соответствующей солености была получена путем добавления аквариумной соли Instant Ocean к фильтрованной морской воде с соленостью окружающей среды залива до достижения целевой солености. Воду в аквариумах меняли раз в неделю. Вторая частичная медуза Aurelia была собрана 3 июля 2017 года в нейстонную сеть и сохранена в 100% этаноле. Оба образца медуз были использованы для молекулярного анализа.
Молекулярное штрих-кодирование для идентификации видов
Полная геномная ДНК была очищена из отдельных полипов, взятых из установленных культур полипов каждого вида, и из тканей двух образцов медуз, консервированных этанолом, собранных на борту R / V Pelican .Субъединица I цитохром с оксидазы митохондрий (COI) и внутренний транскрибируемый спейсер 1 ядра (ITS1) были использованы для характеристики на уровне видов. COI амплифицировали с использованием праймеров LCOjf (Dawson et al., 2005) и HCO2198 (Folmer et al., 1994) с использованием протокола термоциклирования, описанного Piraino et al.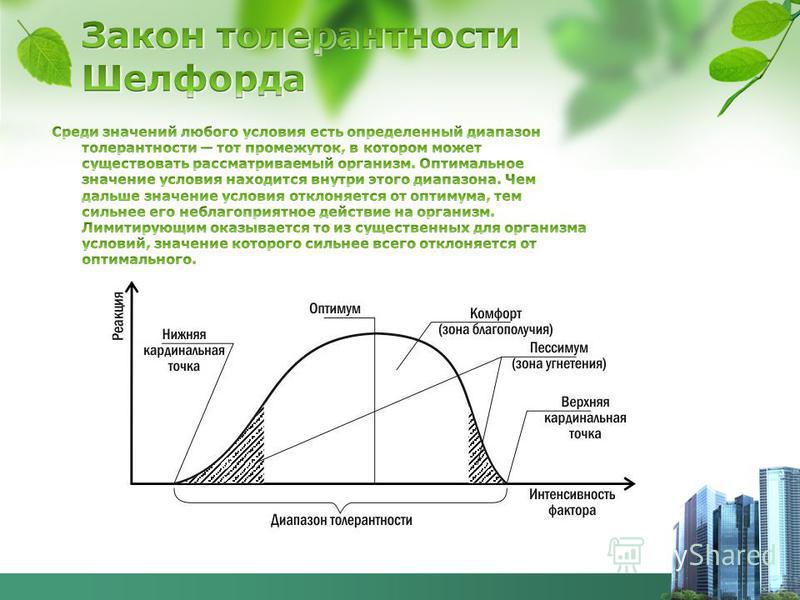 (2014). ITS1 амплифицировали с использованием праймеров KMBN-8 и KMBN-84 от Chiaverano et al. (2016), используя протокол термоциклирования, описанный авторами. Все полимеразные цепные реакции (ПЦР) проводили в термоциклере BioRad.Для проверки качества и размера ампликонов продукты ПЦР визуализировали на 1,5% агарозном геле, окрашенном SYBR Safe. Продукты ПЦР очищали с использованием ExoSAP-IT TM (Applied Biosystems) или набора для экстракции геля GeneJET (Thermo Scientific). Ампликоны COI были двунаправленно секвенированы Core Lab Genomics Corpus Christi Техасского университета с использованием праймеров для ПЦР. Последовательности просматривали и собирали в Geneious 9.1.8. Для идентификации видов каждая согласованная последовательность опрашивалась с использованием алгоритма поиска BLASTn в базе данных коллекции нуклеотидов (nr / nt) Национального центра биотехнологии (NCBI, http: // www.ncbi.nlm.nih.gov).
(2014). ITS1 амплифицировали с использованием праймеров KMBN-8 и KMBN-84 от Chiaverano et al. (2016), используя протокол термоциклирования, описанный авторами. Все полимеразные цепные реакции (ПЦР) проводили в термоциклере BioRad.Для проверки качества и размера ампликонов продукты ПЦР визуализировали на 1,5% агарозном геле, окрашенном SYBR Safe. Продукты ПЦР очищали с использованием ExoSAP-IT TM (Applied Biosystems) или набора для экстракции геля GeneJET (Thermo Scientific). Ампликоны COI были двунаправленно секвенированы Core Lab Genomics Corpus Christi Техасского университета с использованием праймеров для ПЦР. Последовательности просматривали и собирали в Geneious 9.1.8. Для идентификации видов каждая согласованная последовательность опрашивалась с использованием алгоритма поиска BLASTn в базе данных коллекции нуклеотидов (nr / nt) Национального центра биотехнологии (NCBI, http: // www.ncbi.nlm.nih.gov).
Диапазоны и пределы теплового допуска
Мы оценили температурные допуски, используя два разных подхода: метод хронической летальной температуры (CLM) и метод критической температуры (CTM). Оба метода используют динамический подход к определению термостойкости, при котором температура постепенно изменяется, пока не будет достигнута заданная конечная точка. Однако методы различаются по скорости изменения температуры и используемой конечной точке (Beitinger et al., 2000) и, следовательно, оценивают различные аспекты термостойкости.Максимальные и минимальные пределы как в CTM, так и в CLM определяются путем расчета среднего арифметического значений конечных температур среди биологических реплик (Vinagre et al., 2018).
Оба метода используют динамический подход к определению термостойкости, при котором температура постепенно изменяется, пока не будет достигнута заданная конечная точка. Однако методы различаются по скорости изменения температуры и используемой конечной точке (Beitinger et al., 2000) и, следовательно, оценивают различные аспекты термостойкости.Максимальные и минимальные пределы как в CTM, так и в CLM определяются путем расчета среднего арифметического значений конечных температур среди биологических реплик (Vinagre et al., 2018).
CLM использует скорость изменения температуры, которая достаточно медленная, чтобы позволить организмам реакклиматизироваться при каждой новой температуре, и использует смерть в качестве конечной точки (Beitinger et al., 2000). Скорость изменения температуры обычно устанавливается на уровне 1 ° C / день или ниже (Beitinger et al., 2000; Eme and Bennett, 2009). Благодаря включению акклиматизации CLM имеет преимущество перед CTM в том, что он дает более точную оценку фактических термических ограничений вида в природе (Beitinger et al. , 2000). Мы использовали подход CLM для оценки максимальной и минимальной температуры, которую каждый вид Aurelia может выдерживать в дикой природе, в форме хронического летального термического максимума (CLMax) и хронического летального термического минимума (CLMin) для каждого вида. Чтобы получить более подробное представление о том, как каждый вид реагирует на изменение температуры, мы контролировали полипы на наличие признаков стресса через регулярные промежутки времени во время повышения и понижения температуры во время испытаний CLM. Мы использовали морфологию щупалец и реакцию полипа на тактильные стимулы (удары металлическим зондом) для мониторинга стресса и создали систему ранжирования, которую мы назвали «оценкой ответа» (таблица 1).Эта оценка основана на наблюдениях, согласно которым в стандартных условиях культивирования полипы реагируют на внешние стимулы немедленными сокращениями мышц и поддерживают щупальца в вытянутом положении, готовые к кормлению. Оценка реакции использовалась для отслеживания способности каждого вида адаптироваться к увеличению или уменьшению температуры, а также для отслеживания наступления термического стресса, ведущего к смерти.
, 2000). Мы использовали подход CLM для оценки максимальной и минимальной температуры, которую каждый вид Aurelia может выдерживать в дикой природе, в форме хронического летального термического максимума (CLMax) и хронического летального термического минимума (CLMin) для каждого вида. Чтобы получить более подробное представление о том, как каждый вид реагирует на изменение температуры, мы контролировали полипы на наличие признаков стресса через регулярные промежутки времени во время повышения и понижения температуры во время испытаний CLM. Мы использовали морфологию щупалец и реакцию полипа на тактильные стимулы (удары металлическим зондом) для мониторинга стресса и создали систему ранжирования, которую мы назвали «оценкой ответа» (таблица 1).Эта оценка основана на наблюдениях, согласно которым в стандартных условиях культивирования полипы реагируют на внешние стимулы немедленными сокращениями мышц и поддерживают щупальца в вытянутом положении, готовые к кормлению. Оценка реакции использовалась для отслеживания способности каждого вида адаптироваться к увеличению или уменьшению температуры, а также для отслеживания наступления термического стресса, ведущего к смерти.
Таблица 1. Оценка ответа с соответствующей морфологией полипа и степенью реакции щупалец и тела.
CTM — распространенный метод определения пределов термостойкости видов (Bennett et al., 2018), который также использовался для оценки реакции беспозвоночных на изменение климата (Madeira et al., 2012; Vinagre et al., 2016, 2018). , 2019). CTM особенно полезен для более точного определения толерантности между видами (Beitinger et al., 2000) и использовался в этом исследовании для определения различий в верхних пределах термостойкости между конгенерами Aurelia . В CTM температура изменяется с постоянной скоростью до тех пор, пока не будет достигнута заранее заданная сублетальная критическая конечная точка.Критическая конечная точка обычно определяется как нелетальная, но выводящая из строя точка (Lutterschmidt and Hutchison, 1997). Темпы изменения температуры CTM устанавливаются достаточно быстро, чтобы не происходила акклиматизация, но достаточно медленными, чтобы температуру можно было отслеживать, и должны быть стандартизированы, чтобы можно было проводить сравнение между видами (Eme and Bennett, 2009; Bennett et al. , 2018). Мы использовали обычно выбранную скорость изменения температуры 1 ° C / 15 мин (Bennett et al., 2018; Vinagre et al., 2018, 2019).
, 2018). Мы использовали обычно выбранную скорость изменения температуры 1 ° C / 15 мин (Bennett et al., 2018; Vinagre et al., 2018, 2019).
Чтобы оценить соленость, которую полипы Aurelia могут переносить в дикой природе, мы использовали подход хронического изменения солености, аналогичный концепции CLM, который мы называем методом хронической летальной солености (CLSM).Мы отслеживали отдельные полипы по мере постепенного увеличения или уменьшения солености и контролировали их кажущийся уровень стресса через регулярные промежутки времени, используя баллы ответа, до конечной точки. Данные оценки реакции использовались для отслеживания способности каждого вида адаптироваться к изменению солености и для отслеживания наступления соленого стресса, ведущего к смерти. Смерть была обозначена как конечная точка. Медленные темпы изменения переменных окружающей среды позволяют полипам физиологически акклиматизироваться, так что пределы толерантности приблизительно соответствуют тому, какие виды будут терпеть в дикой природе. Мы выбрали скорость изменения солености 1 ppt в день, чтобы максимально увеличить время акклиматизации в практических пределах для исследователей. Используя этот подход, пределы солености были рассчитаны путем взятия среднего арифметического значений солености летальных конечных точек среди биологических повторов (CLSMin и CLSMax).
Мы выбрали скорость изменения солености 1 ppt в день, чтобы максимально увеличить время акклиматизации в практических пределах для исследователей. Используя этот подход, пределы солености были рассчитаны путем взятия среднего арифметического значений солености летальных конечных точек среди биологических повторов (CLSMin и CLSMax).
Диапазон хронической термической акклиматизации и CLM
Для каждого видов Aurelia помещали три полипа, не размножающихся бесполым путем, и позволяли им осесть в каждой лунке 12-луночного планшета для культивирования с 7 мл морской воды при солености 33 ppt.Полипы были выбраны из случайных мест в родительской культуре, чтобы максимизировать генетическое разнообразие и предотвратить выбор полипов, принадлежащих к одной и той же клональной линии. Через 3 дня полипы проверяли на прикрепление, и один здоровый прикрепленный полип оставался в чашке; все остальные были удалены. Для каждого вида использовали три репликационных культуральных планшета, всего 36 биологических повторов (полипов) на вид. Чашки с экспериментальной культурой помещали в инкубатор при выключенном свете.Температуру постепенно повышали, начиная с 21 ° C и увеличивая на 1 ° C в день. 21 ° C была средней температурой условий культивирования в помещении, где хранились многолетние культуры. Эта температура акклиматизации была выбрана для минимизации исходного физиологического стресса полипов перед началом экспериментов по адаптации к хронической температуре. Один 12-луночный культуральный планшет, заполненный 12 полипами / видами, использовали в качестве контроля и выдерживали в инкубаторе при 21 ° C с выключенным светом на время эксперимента.Полипы кормили примерно 10 науплиев A. salina и 15 коловратками на лунку каждое третье утро (каждые 2 ° C) в течение 2–3 часов. После кормления производилась полная подмена воды. Воду для всех культур получали с использованием натуральной фильтрованной морской воды, доведенной до целевой солености 33 ppt с использованием морской соли Instant Ocean. Перед каждой подменой воды в инкубаторах предварительно подогревали воду до заданной температуры.
Чашки с экспериментальной культурой помещали в инкубатор при выключенном свете.Температуру постепенно повышали, начиная с 21 ° C и увеличивая на 1 ° C в день. 21 ° C была средней температурой условий культивирования в помещении, где хранились многолетние культуры. Эта температура акклиматизации была выбрана для минимизации исходного физиологического стресса полипов перед началом экспериментов по адаптации к хронической температуре. Один 12-луночный культуральный планшет, заполненный 12 полипами / видами, использовали в качестве контроля и выдерживали в инкубаторе при 21 ° C с выключенным светом на время эксперимента.Полипы кормили примерно 10 науплиев A. salina и 15 коловратками на лунку каждое третье утро (каждые 2 ° C) в течение 2–3 часов. После кормления производилась полная подмена воды. Воду для всех культур получали с использованием натуральной фильтрованной морской воды, доведенной до целевой солености 33 ppt с использованием морской соли Instant Ocean. Перед каждой подменой воды в инкубаторах предварительно подогревали воду до заданной температуры.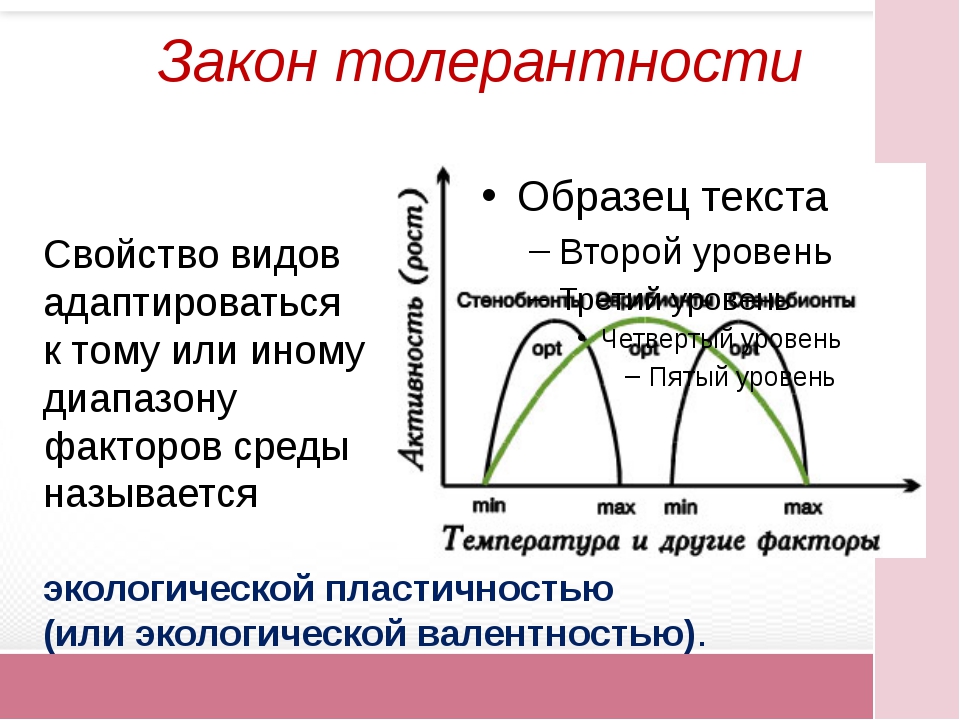 Параметры воды были проверены с помощью YSI для поддержания точности. Температуру внутри инкубаторов регистрировали с помощью HOBO Onset temperature и light logger.Полипы наблюдали под диссекционным микроскопом Leica через каждые 2 ° C, пока их оценка реакции не достигла 3, затем их наблюдали через каждые 1 ° C. Полипы оценивали по признакам стресса по 5-балльной шкале, основанной на морфологии их щупалец и реакции на раздражители (таблица 1). Металлический зонд или пластиковый наконечник пипетки касались щупалец и тела каждого полипа, чтобы оценить реакцию на раздражители (рис. 1). Регистрировали температуру, оценку ответа каждого полипа и количество полипов в каждой лунке.Любые самостоятельные детские полипы или свободно плавающая эфира были удалены. Считалось, что полипы достигли летальной конечной точки, когда они потеряли целостность ткани при оценке реакции 1, которая определялась как отсутствие отдачи ткани при протыкании тела полипа зондом. Затем эксперимент был повторен, но с понижением температуры.
Параметры воды были проверены с помощью YSI для поддержания точности. Температуру внутри инкубаторов регистрировали с помощью HOBO Onset temperature и light logger.Полипы наблюдали под диссекционным микроскопом Leica через каждые 2 ° C, пока их оценка реакции не достигла 3, затем их наблюдали через каждые 1 ° C. Полипы оценивали по признакам стресса по 5-балльной шкале, основанной на морфологии их щупалец и реакции на раздражители (таблица 1). Металлический зонд или пластиковый наконечник пипетки касались щупалец и тела каждого полипа, чтобы оценить реакцию на раздражители (рис. 1). Регистрировали температуру, оценку ответа каждого полипа и количество полипов в каждой лунке.Любые самостоятельные детские полипы или свободно плавающая эфира были удалены. Считалось, что полипы достигли летальной конечной точки, когда они потеряли целостность ткани при оценке реакции 1, которая определялась как отсутствие отдачи ткани при протыкании тела полипа зондом. Затем эксперимент был повторен, но с понижением температуры. Температура постепенно снижалась, начиная с 21 ° C по 1 ° C в день. Полипы культивировали, кормили и контролировали, как описано, на предмет повышения температуры.
Температура постепенно снижалась, начиная с 21 ° C по 1 ° C в день. Полипы культивировали, кормили и контролировали, как описано, на предмет повышения температуры.
Рисунок 1. Примеры оценок ответа для полипов Aurelia sp.1. Ответ на раздражители в порядке убывания слева направо 5-1. Оценка ответа 5 указывает на оптимальный ответ полипа; оценка ответа 1 указывает на нарушение целостности ткани (полное определение оценок ответа см. в таблице 1).
Расчет температурных пределов хронической летальности (CLMax и CLMin)
CLMax для каждого вида определяли путем усреднения температур, при которых каждый полип достигал смертельной конечной точки, с использованием уравнения:
CLMax = вид Σ (T) конечная точка / n
Где конечная точка T — это температура, при которой полипы имели оценку 1 во время испытаний CLMax, а n — размер выборки.CLMin для каждого вида рассчитывали с использованием того же уравнения, но с использованием данных испытаний CLMin.
Критический температурный максимум (CTMax)
От пятидесяти до ста здоровых полипов каждого вида Aurelia были перенесены из разных мест в основных культурах в контейнеры емкостью 700 мл с 33 ppt воды и аэрацией. Каждый вид был помещен в инкубатор и выдержан при 21 ° C в течение 2 недель для акклиматизации. 21 ° C использовалась в качестве температуры акклиматизации, чтобы приблизительно соответствовать средней зимней температуре моря на шельфе северного побережья Австралии (Boyer et al., 2011), где могут располагаться естественные популяции полипов. Полипы кормили ad libitum только что вылупившимися науплиями A. salina два раза в неделю в течение 24 часов. В инкубаторе был выключен свет. Воду в емкостях меняли на следующий день после кормления. Полипы голодали в течение 24 часов перед экспериментами CTMax. 5 полипов на лунку и 10 полипов на вид помещали в пропиленовые лунки диаметром 24,1 мм с размером ячеек 420 мкм (TedPella). Колодцы вставляли в пену так, чтобы они плавали, и помещали в термостабильную водяную баню с интенсивной аэрацией.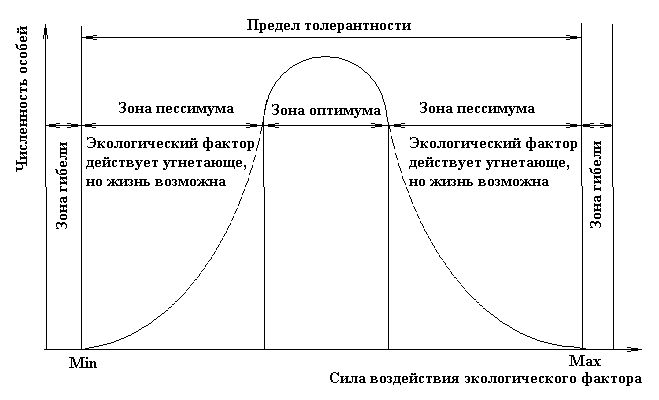 Параметры солености и температуры поддерживались такими же, как при акклиматизации. Полипам давали возможность осесть в течение 24 часов при 21 ° C. Температуру повышали со скоростью 1 ° C / 15 мин. Реакцию каждого полипа на стимулы оценивали при каждом 1 ° C, касаясь щупалец металлическим зондом. Если реакции не наблюдалось, тело полипа касалось металлическим зондом. Ответ на раздражители наблюдали под рассекающим микроскопом Leica из-за небольшого размера полипов. Температуру водяной бани измеряли с помощью цифрового термометра непосредственно перед удалением полипов из водяной бани для наблюдения.Отдельные лунки осторожно зачерпывали водой из водяной бани, используя пластиковый контейнер, и помещали под микроскоп. Когда не наблюдалось никакой реакции на стимулы от щупалец или тела полипа, считалось, что полип достиг своей конечной точки, а температура водяной бани регистрировалась как тепловой максимум полипа. Соленость поддерживалась на уровне 33 ppt в течение всего эксперимента, в то время как концентрация растворенного кислорода и pH контролировались для обеспечения постоянных уровней.
Параметры солености и температуры поддерживались такими же, как при акклиматизации. Полипам давали возможность осесть в течение 24 часов при 21 ° C. Температуру повышали со скоростью 1 ° C / 15 мин. Реакцию каждого полипа на стимулы оценивали при каждом 1 ° C, касаясь щупалец металлическим зондом. Если реакции не наблюдалось, тело полипа касалось металлическим зондом. Ответ на раздражители наблюдали под рассекающим микроскопом Leica из-за небольшого размера полипов. Температуру водяной бани измеряли с помощью цифрового термометра непосредственно перед удалением полипов из водяной бани для наблюдения.Отдельные лунки осторожно зачерпывали водой из водяной бани, используя пластиковый контейнер, и помещали под микроскоп. Когда не наблюдалось никакой реакции на стимулы от щупалец или тела полипа, считалось, что полип достиг своей конечной точки, а температура водяной бани регистрировалась как тепловой максимум полипа. Соленость поддерживалась на уровне 33 ppt в течение всего эксперимента, в то время как концентрация растворенного кислорода и pH контролировались для обеспечения постоянных уровней.
Расчет критического теплового максимума (CTMax)
CTMax для каждого вида рассчитывали путем усреднения температур, при которых полипы теряли реакцию на раздражители, с использованием уравнения:
CTMax = вид Σ (T) конечная точка / n
Где конечная точка T — это температура, при которой полипы теряют реакцию на стимулы, а n — размер выборки. Внутривидовую изменчивость CTMax определяли путем расчета коэффициента вариации, выраженного в процентах для каждого вида, с использованием уравнения: (стандартное отклонение / среднее значение) * 100.
Диапазон акклиматизации к хронической солености и CLSMin
Для каждого видов Aurelia три полипа, не размножающихся бесполым путем, помещали в каждую лунку 12-луночного культурального планшета с 7 мл морской воды с соленостью 33 ppt и давали им прикрепиться. Через три дня полипы проверяли на прикрепление. Один здоровый прикрепленный полип остался в чашке, а остальные удалили. Для каждого вида использовали две копии культуральных пластин, всего 24 полипа на вид.Чашки с культурами помещали в инкубаторы при 19 ° C, что является средней зимней температурой океана на северном побережье GoM (Boyer et al., 2011). Конкретная температура приближается к зимним тепловым условиям эстуариев, заливов и береговой линии региона, где, вероятно, расположены потенциальные места обитания прибрежных полипов. В инкубаторах выключили свет. Соленость была увеличена на 1 ppt в день за счет подмены воды водой соответствующей солености. Восемь полипов каждого вида использовали для контроля и выдерживали в инкубаторе при 19 ° C и солености 33 ppt.Вода для всех культур была приготовлена из натуральной фильтрованной морской воды, доведенной до целевого уровня солености с использованием морской соли Instant Ocean. Воду предварительно нагревали в инкубаторе до 19 ° C, а параметры воды проверяли с помощью YSI перед подменой воды. Температуру внутри инкубатора регистрировали с помощью HOBO Onset temperature и световых логгеров.
Для каждого вида использовали две копии культуральных пластин, всего 24 полипа на вид.Чашки с культурами помещали в инкубаторы при 19 ° C, что является средней зимней температурой океана на северном побережье GoM (Boyer et al., 2011). Конкретная температура приближается к зимним тепловым условиям эстуариев, заливов и береговой линии региона, где, вероятно, расположены потенциальные места обитания прибрежных полипов. В инкубаторах выключили свет. Соленость была увеличена на 1 ppt в день за счет подмены воды водой соответствующей солености. Восемь полипов каждого вида использовали для контроля и выдерживали в инкубаторе при 19 ° C и солености 33 ppt.Вода для всех культур была приготовлена из натуральной фильтрованной морской воды, доведенной до целевого уровня солености с использованием морской соли Instant Ocean. Воду предварительно нагревали в инкубаторе до 19 ° C, а параметры воды проверяли с помощью YSI перед подменой воды. Температуру внутри инкубатора регистрировали с помощью HOBO Onset temperature и световых логгеров. Полипы кормили примерно 10 науплиев A. salina и 15 коловратками на лунку один раз в неделю в течение 2–3 часов. После кормления производилась полная подмена воды.Полипы наблюдали для сбора данных под диссекционным микроскопом Leica при каждом приращении 2 ppt до тех пор, пока их оценка ответа не достигала 3, затем их наблюдали при каждом приращении 1 ppt. Полипы оценивали на предмет видимого уровня стресса в соответствии с 5-балльной шкалой оценки реакции, определенной выше (таблица 1). Металлический зонд или пластиковый наконечник пипетки использовали для прикосновения к щупальцам и телу каждого полипа для оценки реакции на раздражители. Регистрировали соленость, оценку ответа каждого полипа и количество полипов в каждой лунке.Любые самостоятельные детские полипы или свободно плавающая эфира были удалены. Полипы считались умершими, когда они теряли целостность ткани, определяемую как отсутствие отдачи ткани при протыкании тела полипа зондом, с оценкой ответа 1. Эксперимент продолжался до тех пор, пока все полипы не достигли оценки ответа 1.
Полипы кормили примерно 10 науплиев A. salina и 15 коловратками на лунку один раз в неделю в течение 2–3 часов. После кормления производилась полная подмена воды.Полипы наблюдали для сбора данных под диссекционным микроскопом Leica при каждом приращении 2 ppt до тех пор, пока их оценка ответа не достигала 3, затем их наблюдали при каждом приращении 1 ppt. Полипы оценивали на предмет видимого уровня стресса в соответствии с 5-балльной шкалой оценки реакции, определенной выше (таблица 1). Металлический зонд или пластиковый наконечник пипетки использовали для прикосновения к щупальцам и телу каждого полипа для оценки реакции на раздражители. Регистрировали соленость, оценку ответа каждого полипа и количество полипов в каждой лунке.Любые самостоятельные детские полипы или свободно плавающая эфира были удалены. Полипы считались умершими, когда они теряли целостность ткани, определяемую как отсутствие отдачи ткани при протыкании тела полипа зондом, с оценкой ответа 1. Эксперимент продолжался до тех пор, пока все полипы не достигли оценки ответа 1. Эксперимент затем был повторен, но с уменьшением солености. Соленость постепенно снижалась на 1 ppt в день, начиная с 33 ppt. Вода целевой солености была получена путем корректировки естественной фильтрованной морской воды до целевой солености с использованием деионизированной воды.Мониторинг качества воды, кормление и сбор данных выполнялись, как описано для повышения солености.
Эксперимент затем был повторен, но с уменьшением солености. Соленость постепенно снижалась на 1 ppt в день, начиная с 33 ppt. Вода целевой солености была получена путем корректировки естественной фильтрованной морской воды до целевой солености с использованием деионизированной воды.Мониторинг качества воды, кормление и сбор данных выполнялись, как описано для повышения солености.
Расчет пределов хронической летальной солености (CLSMin)
CLSMax не удалось определить, поскольку толерантность полипов превышала диапазон YSI (42 ppt). CLSMin для каждого вида был рассчитан путем усреднения смертельной конечной солености для каждого вида из испытания на уменьшение солености с использованием уравнения:
CLSMin = вид Σ (S) конечная точка / n
Где конечная точка S — это соленость, при которой полипы потеряли целостность ткани при оценке ответа 1, а n — размер выборки.
Результаты
Идентификация с помощью молекулярного штрих-кодирования
Последовательности субъединицы I цитохром с-оксидазы имели длину приблизительно 650 п. н., что является стандартной длиной для Aurelia COI. Последовательности ITS1 составляли около 600 п.н. Было подтверждено, что полипы из прибрежных медуз, собранных в заливе Галвестон, принадлежат к Aurelia sp. 9 согласно COI с процентной идентичностью 99,83% и значением E , равным 0. Полипы, предоставленные аквариумом Moody Gardens, принадлежали A.coerulea на основе COI с процентной идентичностью 100% и E -значением 0. Для морских видов Aurelia наибольшее совпадение BLASTn для митохондриального COI было с A. relicta (номер доступа KX691571), с Процент идентичности 91,33% и значение E , равное 0. Лучшим совпадением для ядерного ITS1 было Aureli a sp. Инчхон с процентом идентичности 84,70% и E — значение 1e-158. Aurelia sp.5 ( A. relicta ) также входила в пятерку лучших совпадений в базе данных и имела более высокий процент идентичности — 88.65% и E — значение 5e-122. Поскольку мы не можем идентифицировать штамм Aurelia с каких-либо известных видов Aurelia , мы называем этот штамм Aurelia sp.
н., что является стандартной длиной для Aurelia COI. Последовательности ITS1 составляли около 600 п.н. Было подтверждено, что полипы из прибрежных медуз, собранных в заливе Галвестон, принадлежат к Aurelia sp. 9 согласно COI с процентной идентичностью 99,83% и значением E , равным 0. Полипы, предоставленные аквариумом Moody Gardens, принадлежали A.coerulea на основе COI с процентной идентичностью 100% и E -значением 0. Для морских видов Aurelia наибольшее совпадение BLASTn для митохондриального COI было с A. relicta (номер доступа KX691571), с Процент идентичности 91,33% и значение E , равное 0. Лучшим совпадением для ядерного ITS1 было Aureli a sp. Инчхон с процентом идентичности 84,70% и E — значение 1e-158. Aurelia sp.5 ( A. relicta ) также входила в пятерку лучших совпадений в базе данных и имела более высокий процент идентичности — 88.65% и E — значение 5e-122. Поскольку мы не можем идентифицировать штамм Aurelia с каких-либо известных видов Aurelia , мы называем этот штамм Aurelia sp. новый. Мультигенный филогенетический анализ для выяснения филогенетического положения A. sp. new внутри рода находится в стадии разработки, но выходит за рамки данной статьи.
новый. Мультигенный филогенетический анализ для выяснения филогенетического положения A. sp. new внутри рода находится в стадии разработки, но выходит за рамки данной статьи.
Пределы хронической термической акклиматизации (CLMin и CLMax)
Полипы A. coerulea сохранили средний балл ответа 5 от 8 ° C до 27 ° C.При температурах выше 27 ° C оценка отклика снизилась с 5 до 1 в течение 3 ° C. При температуре ниже 8 ° C оценка ответа полипов A. coerulea снизилась до 3 при 5 ° C. Полипы A. coerulea не показали видимых изменений в уровне стресса от 5 ° C до 0 ° C, но оценка реакции упала с 3 до 1, когда температура снизилась с 0 ° C до –2 ° C. Потеря реакции на раздражители происходила примерно при –1 ° C (рис. 2). Для полипов A. coerulea CLMax составлял 30,9 ° C, а CLMin составлял –2 ° C; все полипы этого вида достигли конечной точки при одинаковой температуре во время эксперимента с понижением температуры (таблица 2).Температурный диапазон для A. coerulea составляет 32,9 ° C.
coerulea составляет 32,9 ° C.
Рис. 2. Оценка ответа видов Aurelia при изменении температуры на 1 ° C / день. Оценка реакции 1–5 представлена в таблице 1. Температура дана в градусах Цельсия (° C). Планки погрешностей отображают стандартную ошибку. Цвета соответствуют видам Aurelia : красный — Aurelia coerulea , зеленый — A. sp. новый, синий — A. sp. 9.
Таблица 2. Пределы хронической летальной температуры для трех видов Aurelia : минимум хронической летальности (CLMin) и максимум хронической летальности (CLMax).
Полипы Aurelia sp. new получил средний балл 5 от 13 ° C до 27 ° C. При температурах выше 27 ° C оценка отклика снизилась с 5 до 1 в течение 2 ° C. Ниже 13 ° C оценка отклика снижалась до 4 на 10 ° C и стабильно до 1 при 6 ° C. Потеря реакции на раздражители произошла примерно при 7 ° C (рис. 2). Для Aurelia sp.новые полипы, CLMax составлял 30 ° C, а CLMin составлял 6 ° C; все полипы этого вида достигают своих конечных точек при одинаковых температурах (Таблица 2). Температурный диапазон на А. sp. новый — 24 ° C.
Температурный диапазон на А. sp. новый — 24 ° C.
Полипы Aurelia sp. 9 поддержали средний балл 5 от 14 ° C до 30 ° C. При температурах выше 14 ° C оценка реакции полипов снижалась с 5 до 1 в течение 5 ° C. Ниже 14 ° C оценка отклика снизилась до 4 при 10 ° C и до 1 при 3 ° C. Потеря ответа на раздражители произошла примерно при 5 ° C (рис. 2).Для Aurelia sp. 9 полипов, CLMax составлял 34,7 ° C, а CLMin составлял 3,1 ° C (таблица 2). Температурный диапазон на А. sp. 9 — 31,6 ° C.
полипов в контрольных группах сохраняли оценку ответа 4 или выше на протяжении всего эксперимента. Результаты для контрольных полипов можно найти в дополнительном материале S1.
Критический температурный максимум (CTMax)
Полипы Aurelia sp. 9 имел самый высокий термостойкость с CTMax 37,2 ° C. Первый полип потерял реакцию на стимулы при 34 ° C, но большинство полипов потеряли реакцию на стимулы выше 37 ° C, причем наиболее толерантные полипы сохраняли реакцию до 38. 4 ° С. Внутривидовая изменчивость для Aurelia sp. 9 было 4,0%. Aurelia sp. новые полипы имели CTMax 32,1 ° C с 0% внутривидовой вариабельностью (все теряли реакцию на стимулы в одно и то же время и при той же температуре). Полипы A. coerulea были наименее устойчивыми к высоким температурам с CTMax 29,6 ° C. Первые полипы теряли реакцию на раздражители при 28,7 ° C. Один полип переносил температуру до 31,8 ° C. Внутривидовая изменчивость для A. coerulea составила 4.1%. Результаты экспериментов CTMax приведены в таблице 3.
4 ° С. Внутривидовая изменчивость для Aurelia sp. 9 было 4,0%. Aurelia sp. новые полипы имели CTMax 32,1 ° C с 0% внутривидовой вариабельностью (все теряли реакцию на стимулы в одно и то же время и при той же температуре). Полипы A. coerulea были наименее устойчивыми к высоким температурам с CTMax 29,6 ° C. Первые полипы теряли реакцию на раздражители при 28,7 ° C. Один полип переносил температуру до 31,8 ° C. Внутривидовая изменчивость для A. coerulea составила 4.1%. Результаты экспериментов CTMax приведены в таблице 3.
Таблица 3. Значения критического теплового максимума (CTMax) для трех видов Aurelia .
Диапазон акклиматизации к солености и CLSMin
Полипы Aurelia coerulea сохранили средний балл ответа от 5 до примерно 12 ppt, с оценкой ответа ниже 4 при примерно 11 ppt. Полипы потеряли реакцию на стимулы примерно на 7 ppt и целостность ткани на 6 ppt. CLSMin для A.coerulea составила 6,2 п. п. Aurelia sp. новые поддерживаемые оценки ответа 5, без признаков видимого стресса, вплоть до солености 18 ppt, и оценки ответа от 4 или выше до 15 ppt. Полипы потеряли реакцию на стимулы примерно на 12 ppt и целостность ткани на 10 ppt. CLSMin для Aurelia sp. новых было 10 п.п. Aurelia sp. 9 полипов сохранили оценку ответа 5 при 10 ppt и оценку ответа от 4 или выше до приблизительно 7 ppt. Полипы теряли реакцию на стимулы примерно на 4 ppt и теряли целостность ткани на 2 ppt.CLSMin для Aurelia sp. 9 составила 2,2 п.п. Значения CLSMin суммированы в таблице 4. В эксперименте по акклиматизации к хронической солености с увеличением солености полипы всех трех видов сохраняли оптимальную оценку реакции до солености 42 ppt, что было пределом измерения для солемера YSI. Верхний предел акклиматизации для солености и CLSMax не может быть определен, но превышает экологически значимые значения для GoM. Полипы в контрольных группах сохраняли средний балл ответа 4 или выше в течение всего эксперимента.
п. Aurelia sp. новые поддерживаемые оценки ответа 5, без признаков видимого стресса, вплоть до солености 18 ppt, и оценки ответа от 4 или выше до 15 ppt. Полипы потеряли реакцию на стимулы примерно на 12 ppt и целостность ткани на 10 ppt. CLSMin для Aurelia sp. новых было 10 п.п. Aurelia sp. 9 полипов сохранили оценку ответа 5 при 10 ppt и оценку ответа от 4 или выше до приблизительно 7 ppt. Полипы теряли реакцию на стимулы примерно на 4 ppt и теряли целостность ткани на 2 ppt.CLSMin для Aurelia sp. 9 составила 2,2 п.п. Значения CLSMin суммированы в таблице 4. В эксперименте по акклиматизации к хронической солености с увеличением солености полипы всех трех видов сохраняли оптимальную оценку реакции до солености 42 ppt, что было пределом измерения для солемера YSI. Верхний предел акклиматизации для солености и CLSMax не может быть определен, но превышает экологически значимые значения для GoM. Полипы в контрольных группах сохраняли средний балл ответа 4 или выше в течение всего эксперимента. Результаты для контрольных полипов можно найти в дополнительном материале S1.
Результаты для контрольных полипов можно найти в дополнительном материале S1.
Таблица 4. Минимум хронической летальной солености (CLSMin) для трех видов Aurelia .
Обсуждение
Изменения окружающей среды, связанные с изменением климата, приводят к изменению ареала видов. Общие тенденции во всем мире показывают, что виды реагируют на повышение температуры океана, сдвигаясь в сторону полюсов (Thomas, 2010). Однако в Правительстве Молдовы Североамериканский континент образует физический барьер, ограничивающий движение видов на север.Из-за сложного жизненного цикла, успех видов медуз зависит от способности полипов, эфиров и медуз на разных этапах жизни переносить будущие условия. Поскольку именно стадия полипа отвечает за поддержание и расширение популяций медуз между сезонами и годами, на успех вида влияет способность полипа переносить повышение температуры.
Морской вид Aurelia может быть новым видом, поскольку он не был представлен среди последовательностей GenBank COI или ITS1. Для подтверждения идентичности Aurelia sp. Требуются дополнительные молекулярные и морфологические анализы. новый как отдельный вид. Только два вида Aurelia были ранее зарегистрированы в Правительстве Молдовы: A. sp. 9 и A. c.f. sp. 2 (Chiaverano et al., 2016).
Для подтверждения идентичности Aurelia sp. Требуются дополнительные молекулярные и морфологические анализы. новый как отдельный вид. Только два вида Aurelia были ранее зарегистрированы в Правительстве Молдовы: A. sp. 9 и A. c.f. sp. 2 (Chiaverano et al., 2016).
Aurelia coerulea , Aurelia sp. новый, и Aurelia sp. 9 имеют четкие диапазоны температурных допусков и тепловые пределы (Таблица 2). В частности, термальные ареалы инвазивных видов A.coerulea и Aurelia sp. 9 имеют одинаковый размер, но отличаются только на 1,3 ° C по пределам летальности. Показатели реакции обоих видов демонстрируют схожую тенденцию, поскольку оба сохранили 5 баллов при температуре 21 ° C (рис. 2). Однако A. coerulea имеет более низкую термостойкость, а его экспериментальный диапазон термостойкости смещен примерно на 4 ° C по сравнению с Aurelia sp. 9, что свидетельствует о предпочтении более низкой температуры. Это также отражено в климате его естественного географического происхождения в Южном и Восточно-Китайском морях.Тепловой диапазон Aurelia sp. new примерно на 8 ° C уже, чем у A. coerulea и A. sp. 9 (рис. 2), предполагая, что полипы этого вида могут предпочитать термически стабильные условия. Значения CLMax и видимый стресс, измеренный по шкале ответов, позволяют предположить, что A. coerulea и A. sp. Новый может иметь аналогичные верхние пределы температуры. Контрольные полипы, которые содержались при постоянной температуре, но в остальном обрабатывались идентичным образом, сохраняли высокие показатели ответа в течение всего периода испытаний, указывая на то, что наблюдаемые пределы летальности были вызваны термическим стрессом.
Это также отражено в климате его естественного географического происхождения в Южном и Восточно-Китайском морях.Тепловой диапазон Aurelia sp. new примерно на 8 ° C уже, чем у A. coerulea и A. sp. 9 (рис. 2), предполагая, что полипы этого вида могут предпочитать термически стабильные условия. Значения CLMax и видимый стресс, измеренный по шкале ответов, позволяют предположить, что A. coerulea и A. sp. Новый может иметь аналогичные верхние пределы температуры. Контрольные полипы, которые содержались при постоянной температуре, но в остальном обрабатывались идентичным образом, сохраняли высокие показатели ответа в течение всего периода испытаний, указывая на то, что наблюдаемые пределы летальности были вызваны термическим стрессом.
В отличие от Aurelia sp. 9 и sp. new, полипы A. coerulea были обнаружены в дикой природе (Ishii, Katsukoshi, 2010; Marques et al., 2019). Интересно, что экспериментально определенный диапазон температур, в котором полипы A. coerulea сохраняли минимальные признаки стресса (оценка реакции 4 или выше), определенный в этом исследовании, хорошо согласуется с опубликованными отчетами о температурах среды обитания для этого вида, полученными в результате полевых исследований диких животных. населения. Согласно нашим экспериментам, А.coerulea подвергались минимальному стрессу от 6,5 ° C до 27 ° C, тогда как естественная среда обитания полипов в лагуне Тау (северо-западное Средиземноморье) колеблется от 7,6 ° C до 25,8 ° C (Marques et al., 2019), от 6 ° C до 30 ° C. ° C для полипов в озере Верано, Италия (Belmonte et al., 2011), и от 9 ° C до 29 ° C для A. coerulea в Токийском заливе, Япония (Ishii and Katsukoshi, 2010). Это минимальная и максимальная зарегистрированная температура воды диких популяций, обследованных в течение примерно 1 года (Ishii and Katsukoshi, 2010; Marques et al., 2019).
coerulea сохраняли минимальные признаки стресса (оценка реакции 4 или выше), определенный в этом исследовании, хорошо согласуется с опубликованными отчетами о температурах среды обитания для этого вида, полученными в результате полевых исследований диких животных. населения. Согласно нашим экспериментам, А.coerulea подвергались минимальному стрессу от 6,5 ° C до 27 ° C, тогда как естественная среда обитания полипов в лагуне Тау (северо-западное Средиземноморье) колеблется от 7,6 ° C до 25,8 ° C (Marques et al., 2019), от 6 ° C до 30 ° C. ° C для полипов в озере Верано, Италия (Belmonte et al., 2011), и от 9 ° C до 29 ° C для A. coerulea в Токийском заливе, Япония (Ishii and Katsukoshi, 2010). Это минимальная и максимальная зарегистрированная температура воды диких популяций, обследованных в течение примерно 1 года (Ishii and Katsukoshi, 2010; Marques et al., 2019).
Температурный диапазон A. coerulea предполагает, что маловероятно развитие постоянной популяции в прибрежном правительстве страны. Низкие зимние температуры вдоль северо-западного и северо-восточного побережий Правительства Минска в среднем 13–20 ° C (по данным https://www.nodc.noaa.gov/dsdt/cwtg/all_meanT.html), что значительно превышает нижний предел термостойкости. предел –2 ° C (CLMin) для A. coerulea. Однако при CLMax 30,9 ° C этот вид может быть ограничен летними температурами воды на некоторых участках северо-западного и северо-восточного побережья Правительства Москвы, которые в среднем составляют 28–31 ° C (среднее значение по https: // www.nodc.noaa.gov/dsdt/cwtg/all_meanT.html). Таким образом, средние летние температуры в прибрежном GoM, вероятно, будут смертельными до A. coerulea , оценка реакции которой указывает на разрушительные уровни теплового стресса при температурах выше 27 ° C.
Aurelia sp. 9, обычное цветение, образующее видов Aurelia в Правительстве штата Мичиган, имело диапазон термической устойчивости от 3,1 ° C до 34,7 ° C, исходя из летальных пределов, и проявляло минимальные признаки стресса между 10 ° C и 31 ° C. Среднемесячные среднегодовые значения для прибрежных западных и восточных районов Мичигана колеблются от 13 ° C до 31 ° C, что свидетельствует о том, что с тепловой точки зрения условия заливов, пристаней для яхт и береговых линий являются подходящими средами обитания для Aurelia sp.9 полипов. Из трех изученных видов Aurelia sp. 9 был устойчив к температурам как минимум на 3 ° C выше, чем два других вида. Aurelia sp. new имел самый узкий температурный диапазон 24 ° C и демонстрировал признаки напряжения за пределами диапазона от 10 ° C до 27 ° C. Верхний предел летальности для Aurelia sp. new (CLMax = 30 ° C) предполагает, что он также может не выдержать высоких летних температур, наблюдаемых вдоль северного побережья Правительства Москвы. Однако температуры в более глубоких водах вдоль континентального шельфа в Правительстве Моря, где были собраны медузы этого вида, обычно ниже, чем средние летние прибрежные значения, поэтому возможно, что Aurelia sp.
Среднемесячные среднегодовые значения для прибрежных западных и восточных районов Мичигана колеблются от 13 ° C до 31 ° C, что свидетельствует о том, что с тепловой точки зрения условия заливов, пристаней для яхт и береговых линий являются подходящими средами обитания для Aurelia sp.9 полипов. Из трех изученных видов Aurelia sp. 9 был устойчив к температурам как минимум на 3 ° C выше, чем два других вида. Aurelia sp. new имел самый узкий температурный диапазон 24 ° C и демонстрировал признаки напряжения за пределами диапазона от 10 ° C до 27 ° C. Верхний предел летальности для Aurelia sp. new (CLMax = 30 ° C) предполагает, что он также может не выдержать высоких летних температур, наблюдаемых вдоль северного побережья Правительства Москвы. Однако температуры в более глубоких водах вдоль континентального шельфа в Правительстве Моря, где были собраны медузы этого вида, обычно ниже, чем средние летние прибрежные значения, поэтому возможно, что Aurelia sp.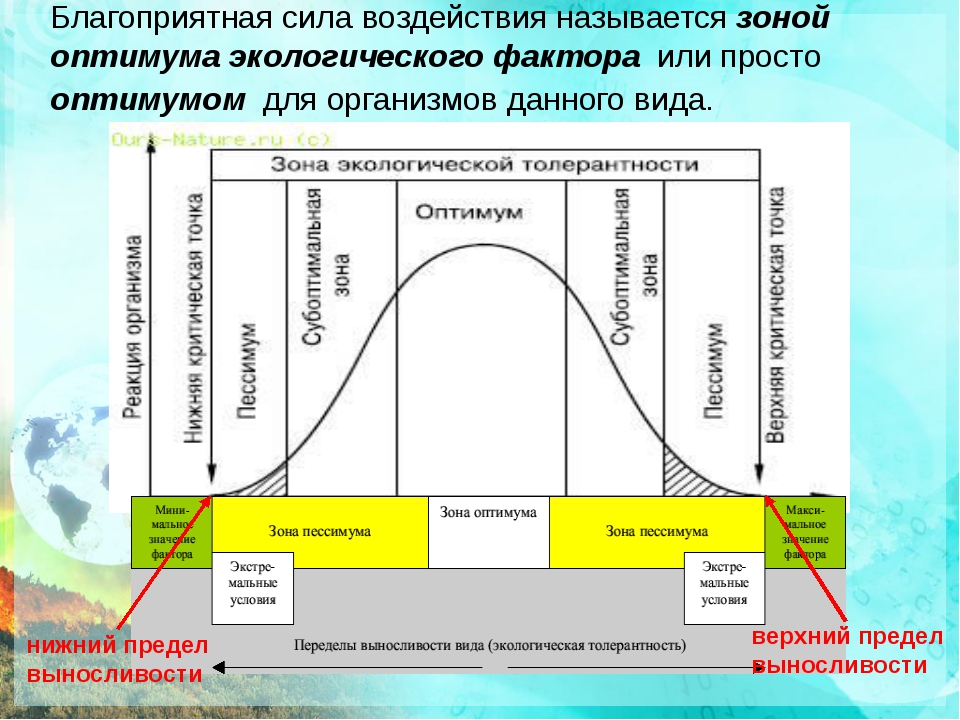 new ограничен офшорными зонами.
new ограничен офшорными зонами.
Мы использовали CTM для определения CTMax для каждого вида, чтобы определить относительные верхние термические пределы для конгенеров Aurelia (Таблица 3). Значения CTMax подтверждают, что Aurelia sp. 9 более устойчив к высоким температурам, чем A. coerulea и Aurelia sp. новый. A. coerulea имеет самый низкий CTMax среди сородичей, что указывает на то, что этот вид наименее устойчив к высоким температурам, что является разумным, учитывая в целом более низкие температуры его естественного диапазона по сравнению с GoM.Быстрые скорости потепления, такие как скорость 1 ° C / 15 мин, использованная в этом исследовании для определения CTMax, могут переоценить фактические верхние пределы термостойкости организмов в природе (Peck et al., 2009). Значения CTMax обычно превышают значения CLMax, потому что более медленные темпы потепления в хроническом эксперименте позволяют больше времени при каждой температуре для накопления и наступления смертельных физиологических эффектов. Эта взаимосвязь наблюдалась для Aurelia sp. 9 и Aurelia sp. new, где значения CTMax были примерно на 2 ° C больше, чем значения CLMax, но не для A.coerulea , где CTMax был примерно на 1 ° C ниже, чем CLMax (рис. 4). Более низкий CTMax может указывать на то, что полипы A. coerulea чувствительны к быстрым изменениям температуры.
Эта взаимосвязь наблюдалась для Aurelia sp. 9 и Aurelia sp. new, где значения CTMax были примерно на 2 ° C больше, чем значения CLMax, но не для A.coerulea , где CTMax был примерно на 1 ° C ниже, чем CLMax (рис. 4). Более низкий CTMax может указывать на то, что полипы A. coerulea чувствительны к быстрым изменениям температуры.
Эксперименты с хроническим засолением показывают, что полипы всех трех видов обычно способны выдерживать широкий диапазон солености и нечувствительны к гиперсоленым условиям (рис. 3). Из трех видов Aurelia sp. 9, имел самое низкое значение CLSMin — 2,2 п.п. Кроме того, полипы этого вида не имели видимых признаков стресса во время снижения солености на 1 ppt / день до тех пор, пока соленость не упала ниже 10 ppt, что позволяет предположить, что A. sp. 9 полипов толерантны к низкой солености и ее изменению. Если рассматривать вместе с высокой термостойкостью этого вида, результаты толерантности к солености позволяют предположить, что прибрежные районы Правительства Молдовы, а также некоторые заливы и эстуарии в этом регионе являются подходящими средами обитания для полипов A. sp. 9. полипы A. coerulea имели CLSMin 6,2 ppt без видимых признаков стресса до тех пор, пока соленость не снизилась ниже 14 ppt. Большинство диких популяций A. coerulea было зарегистрировано при солености выше 20 ppt (Belmonte et al., 2011; Marques et al., 2019). Aurelia sp. new имел CLSMin 10 ppt и поэтому был наименее устойчивым к низкой солености среди трех видов. Видимые признаки стресса наблюдались у полипов этого вида при солености ниже 18 ppt, что свидетельствует о возможной низкой устойчивости к изменению солености (Таблица 4). По сравнению с двумя прибрежными видами Aurelia sp. new имеет значительно более ограниченную способность противостоять изменениям температуры и солености, что указывает на A. sp. new — это офшорный вид в Правительстве Молдовы.
Рис. 3. Оценка реакции видов Aurelia в ответ на изменение солености на 1 ppt / день. Оценки ответов 1–5 представлены в таблице 1. Соленость выражена в частях на тысячу (ppt).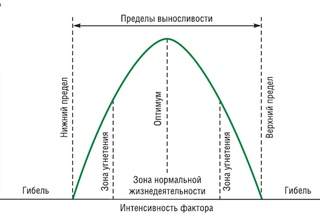 Планки погрешностей отображают стандартную ошибку. Цвета соответствуют видам Aurelia : красный — Aurelia coerulea , зеленый — A. sp. новый, синий — A. sp. 9.
Планки погрешностей отображают стандартную ошибку. Цвета соответствуют видам Aurelia : красный — Aurelia coerulea , зеленый — A. sp. новый, синий — A. sp. 9.
Рисунок 4. Сравнение значений термостойкости в экспериментах с хроническим летальным максимумом и минимумом и критическим термическим максимумом для Aurelia coerulea, A. sp. новый, и A. sp. 9. Температурные пределы указаны в градусах Цельсия (° C). Цвета обозначают тип теплового ограничения: красный — хронический смертельный максимум (CLMax), зеленый — хронический смертельный минимум (CLMin), синий — критический тепловой максимум (CTMax). Коробчатые диаграммы отображают медианное значение с нижним и верхним шарнирами, соответствующими 25-му и 75-му перцентилям, а усы — до 1.В 5 раз больше межквартильного диапазона. Отдельные точки указывают на выбросы.
Из-за трудностей обнаружения полипов в дикой природе мы признаем, что не можем контролировать или оценивать закономерности родства в полипах, используемых в наших экспериментах. Необходимы дополнительные молекулярные, морфологические и физиологические исследования, чтобы подтвердить полученные здесь результаты толерантности к окружающей среде, особенно для A. sp. new, где полипы были получены от одной медузы и могут неадекватно отражать генетическую изменчивость вида.
Необходимы дополнительные молекулярные, морфологические и физиологические исследования, чтобы подтвердить полученные здесь результаты толерантности к окружающей среде, особенно для A. sp. new, где полипы были получены от одной медузы и могут неадекватно отражать генетическую изменчивость вида.
Диапазоны и пределы толерантности к температуре и солености для представителей рода Aurelia можно использовать для прогнозирования реакции видов на будущие условия. Согласно прогнозам, к 2100 году температура поверхности океана вырастет на 4 ° C (Biasutti et al., 2012), а бентические среды обитания станут еще теплее (Turner et al., 2017). Повышение температуры такой величины может удержать A. coerulea от вторжения или укоренения в прибрежном GoM, поскольку верхний тепловой предел этого вида уже равен или ниже текущих средних летних температур воды.Высокие температуры в Южном и Восточно-Китайском морях (26–29 ° C) предполагают, что A. coerulea в настоящее время обитает довольно близко к своим тепловым пределам в своем естественном ареале и может подвергаться особому риску в закрытых средах обитания, что известно обитать. Из-за аналогичных верхних температурных пределов Aurelia sp. новые также могут иметь негативное влияние в целом. Верхний предел хронической летальной температуры (34,7 ° C) и критический температурный максимум (37,2 ° C) Aurelia sp. 9 предполагают, что он может выдерживать некоторое повышение температуры окружающей среды.Однако повышение температуры на 4 ° C приблизит температуру воды к верхнему пределу температуры для этого вида, что потенциально может отрицательно повлиять на Aurelia sp. 9 в самых теплых крайностях своего биогеографического диапазона. Ожидается, что повышение температуры будет наиболее значительным в прибрежных районах (Biasutti et al., 2012), что указывает на то, что прибрежные зоны Aurelia sp. 9 популяций не выиграют и могут сократиться из-за повышения температуры среды обитания к следующему столетию.
Из-за аналогичных верхних температурных пределов Aurelia sp. новые также могут иметь негативное влияние в целом. Верхний предел хронической летальной температуры (34,7 ° C) и критический температурный максимум (37,2 ° C) Aurelia sp. 9 предполагают, что он может выдерживать некоторое повышение температуры окружающей среды.Однако повышение температуры на 4 ° C приблизит температуру воды к верхнему пределу температуры для этого вида, что потенциально может отрицательно повлиять на Aurelia sp. 9 в самых теплых крайностях своего биогеографического диапазона. Ожидается, что повышение температуры будет наиболее значительным в прибрежных районах (Biasutti et al., 2012), что указывает на то, что прибрежные зоны Aurelia sp. 9 популяций не выиграют и могут сократиться из-за повышения температуры среды обитания к следующему столетию.
Заявление о доступности данных
Наборы данных последовательностей ДНК для этого исследования депонированы в GenBank, номера доступа от MN531714 до MN531721 для COI и от MN527964 до MN527966 для ITS1. Генетические данные будут доступны через Центр информации и данных исследовательской инициативы Мексиканского залива (GRIIDC). Данные о допуске по температуре и солености доступны в дополнительном материале S2.
Генетические данные будут доступны через Центр информации и данных исследовательской инициативы Мексиканского залива (GRIIDC). Данные о допуске по температуре и солености доступны в дополнительном материале S2.
Авторские взносы
AF и MM разработали эксперименты и написали рукопись. AF проводил эксперименты.
Финансирование
Это исследование было поддержано грантом Texas Sea № 02-S170210, предоставленным MM, и грантом Texas Sea Grant-in-Aid для AF. Отбор проб был проведен во время исследовательского рейса, проводимого Лабораторией экологии рыболовства Техасского университета A&M при поддержке гранта Исследовательской инициативы Мексиканского залива (Deep Pelagic Nekton Dynamics Consortium).
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим доктора Лауру Юргенс за ее предложения по экспериментальному дизайну. Мы благодарим команду аквариумистов Moody Gardens за предоставленные полипы Aurelia . Мы также благодарим Кэти Сент-Клер и персонал центра морской жизни ТАМУГ за их опыт и поддержку в создании культур медуз и уходе за медузами на этапах их жизни, а также Бетсабе Эррера за ее помощь в ежедневном содержании медуз.Мы выражаем благодарность доктору Джею Р. Рукеру и Лаборатории экологии рыболовства Техасского университета A&M за возможность сопровождать их исследовательский круиз на борту R / V Pelican компании LUMCON. Мы также хотели бы поблагодарить команду R / V Pelican за их помощь в сборе и обнаружении медуз.
Мы благодарим команду аквариумистов Moody Gardens за предоставленные полипы Aurelia . Мы также благодарим Кэти Сент-Клер и персонал центра морской жизни ТАМУГ за их опыт и поддержку в создании культур медуз и уходе за медузами на этапах их жизни, а также Бетсабе Эррера за ее помощь в ежедневном содержании медуз.Мы выражаем благодарность доктору Джею Р. Рукеру и Лаборатории экологии рыболовства Техасского университета A&M за возможность сопровождать их исследовательский круиз на борту R / V Pelican компании LUMCON. Мы также хотели бы поблагодарить команду R / V Pelican за их помощь в сборе и обнаружении медуз.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmars.2020.00093 / полный # дополнительный материал
МАТЕРИАЛ S1 | Ежедневные усредненные оценки ответа для экспериментов с максимальным и минимальным хроническим летальным исходом для контрольных и экспериментальных полипов. Вкладка CLM_controls содержит среднесуточную оценку ответа для контролей каждого вида, поддерживаемую при 21 ° C.
Вкладка CLM_controls содержит среднесуточную оценку ответа для контролей каждого вида, поддерживаемую при 21 ° C.
МАТЕРИАЛ S2 | Необработанные и обработанные данные для экспериментов по устойчивости к температуре и солености. Вкладка CLMin и CLMax содержит значения Chronic Lethal Minimum (CLMin) и Chronic Lethal Maximum (CLMax) для трех видов с указанием размера выборки, стандартного отклонения и размера выборки (CLMax — CLMin).Значения CLMax и CLMin указаны в градусах Цельсия (° C). Вкладка CTMax содержит значения CTMax для трех видов с указанием размера выборки, стандартного отклонения и внутривидовой изменчивости. Значения CTMax указаны в градусах Цельсия (° C). Внутривидовая изменчивость показана в процентах. Вкладка CLSMin содержит значения хронической летальной минимальной солености для трех видов с указанием размера выборки и стандартного отклонения. Значения CLSMin выражены в частях на тысячу (ppt). Вкладка Response Score (температура) содержит исходные данные для экспериментов с хронической летальной температурой.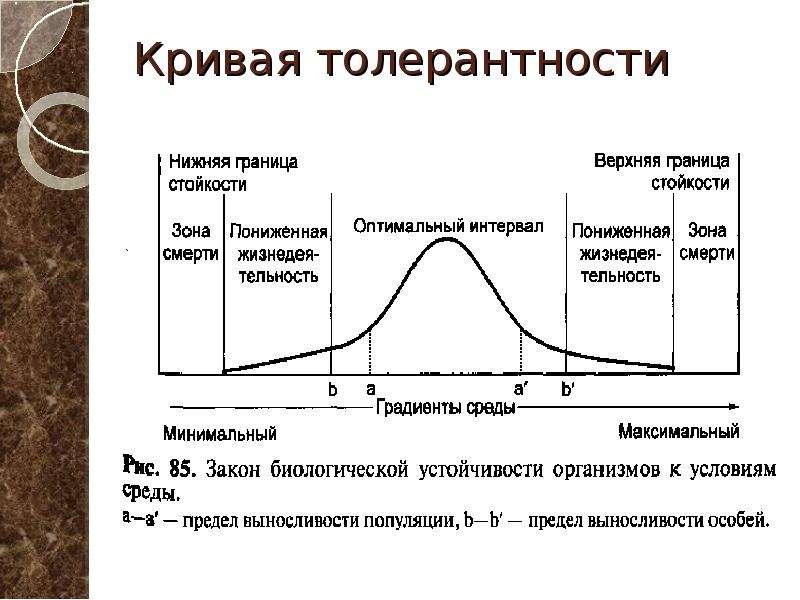 Отображаются дата, температура в градусах Цельсия (° C), вид, номер планшета, номер лунки, оценка ответа и общее количество полипов в лунке. Вкладка Response Score (соленость) содержит исходные данные для экспериментов с хронической летальной соленостью. Отображаются дата, температура в градусах Цельсия (° C), вид, номер планшета, номер лунки, оценка ответа и общее количество полипов в лунке.
Отображаются дата, температура в градусах Цельсия (° C), вид, номер планшета, номер лунки, оценка ответа и общее количество полипов в лунке. Вкладка Response Score (соленость) содержит исходные данные для экспериментов с хронической летальной соленостью. Отображаются дата, температура в градусах Цельсия (° C), вид, номер планшета, номер лунки, оценка ответа и общее количество полипов в лунке.
Список литературы
Абдул Азис, П. К., Ат-Тисан, И., Аль-Дайли, М., Грин, Т. Н., Далви, А. Г. И., и Джавид, М.А. (2000). Воздействие окружающей среды на источник воды для опреснительных заводов на восточном побережье Саудовской Аравии. Опреснение 132, 29–40. DOI: 10.1016 / s0011-9164 (00) 00132-6
CrossRef Полный текст | Google Scholar
Байтингер, Т. Л., Беннет, В. А., и МакКоли, Р. В. (2000). Температурная устойчивость пресноводных рыб Северной Америки, подверженных динамическим изменениям температуры. Environ. Биол. Рыбы. 58, 237–275.
Google Scholar
Бельмонте, Г., Сирокко, Т., и Денитто, Ф. (2011). Состав зоопланктона озера Варано (побережье Адриатического моря, Италия). Ital. J. Zool. 78, 370–378. DOI: 10.1080 / 11250003.2011.561261
CrossRef Полный текст | Google Scholar
Беннетт, Дж. М., Калози, П., Клюзелла-Труллас, С., Мартинес, Б., Сандей, Дж., И Алгар, А. С. (2018). GlobTherm, глобальная база данных по термостойкости для водных и наземных организмов. Sci. Данные 5: 180022. DOI: 10.1038 / sdata.2018.22
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биазутти, М., Собель, А. Х., Камарго, С. Дж., И Крейтс, Т. Т. (2012). Прогнозируемые изменения физического климата на побережье Мексиканского залива и Карибского бассейна. Клим. Чанг. 112, 819–845. DOI: 10.1007 / s10584-011-0254-y
CrossRef Полный текст | Google Scholar
Бойер, Т. П., Биддл, М., Гамильтон, М., Мишонов, А. В., Пейвер, К. Р., Сеидов, Д. и др. (2011). Региональная климатология Мексиканского залива NOAA / NODC. Серебряный источник: NOAA.
Google Scholar
Ceh, J., Гонсалес, Дж., Пачеко, А. С., и Риаскос, Дж. М. (2015). Неуловимый жизненный цикл сцифозных медуз — новый взгляд на метагенез. Sci. Rep. 5, 12037–12037. DOI: 10.1038 / srep12037
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кьяверано, Л. М., Байха, К. В., и Грэм, В. М. (2016). Сравнение локальных и генерализованных фенотипов у двух симпатрических видов Aurelia : понимание экологии медуз с использованием генетики и морфометрии. PLoS One 11: e0156588.DOI: 10.1371 / journal.pone.0156588
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кондон Р. Х., Лукас К. Х., Питт К. А. и Уай С. И. (2014). Цветение медузы и экологические взаимодействия. Mar. Ecol. Прог. Сер. 510, 109–110.
Google Scholar
Доусон, М. Н., Гупта, А. С., и Англия, М. Х. (2005). Сочетание биофизической модели глобального океана и молекулярно-генетического анализа позволяет выявить множественные интродукции криптогенных видов. Proc. Natl. Акад. Sci. США 102, 11968–11973. DOI: 10.1073 / pnas.0503811102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Доусон, М. Н., и Мартин, Л. Е. (2001). Географическая изменчивость и экологическая адаптация Aurelia (Scyphozoa, Semaeostomeae): некоторые выводы из молекулярной филогенетики. Hydrobiologia 451, 259–273. DOI: 10.1007 / 978-94-010-0722-1_21
CrossRef Полный текст | Google Scholar
Де Донно, А., Идоло, А., Багордо, Ф., Грасси, Т., Леоманни, А., и Серио, Ф. (2014). Влияние распространения жалящих медуз на южных побережьях Италии: опасность для здоровья человека, лечение и социальные издержки. Внутр. J. Environ. Res. Общественное здравоохранение 11, 2488–2503. DOI: 10.3390 / ijerph210302488
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дуарте, К. М., Питт, К. А., Лукас, К. Х., Перселл, Дж. Э., Уай, С.-И., и Робинсон, К. (2013). Является ли глобальное разрастание океана причиной цветения медуз? Фронт.Ecol. Environ. 11, 91–97. DOI: 10.1890 / 110246
CrossRef Полный текст | Google Scholar
Эме Дж. И Беннетт У. А. (2009). Критические полигоны термостойкости тропических морских рыб из Сулавеси, Индонезия. J. Therm. Биол. 34, 220–225. DOI: 10.1016 / j.jtherbio.2009.02.005
CrossRef Полный текст | Google Scholar
Фольмер, О., Блэк, М., Хое, В., Лутц, Р., и Вриенхук, Р. (1994). Праймеры ДНК для амплификации субъединицы I митохондриальной цитохром с оксидазы из. Мол. Mar. Biol. Biotechnol. 3, 294–299.
Google Scholar
Грэм, В. М. (2001). Численное увеличение и сдвиги в распределении Chrysaora quinquecirrha (Desor) и Aurelia aurita (Linné) (Cnidaria: Scyphozoa) в северной части Мексиканского залива. Hydrobiologia 451, 97–111. DOI: 10.1007 / 978-94-010-0722-1_9
CrossRef Полный текст | Google Scholar
Gröndahl, F. (1988). Сравнительное экологическое исследование сцифозов Aurelia aurita , Cyanea capillata и C.lamarckii в Гульмар-фьорде, запад Швеции, с 1982 по 1986 год. Mar. Biol. 97, 541–550. DOI: 10.1007 / bf003
CrossRef Полный текст | Google Scholar
Харальдссон, М., Тённессон, К., Тизелиус, П., Тингстад, Т. Ф., и Акснес, Д. Л. (2012). Взаимоотношения между рыбами и медузами в зависимости от эвтрофикации и прозрачности воды. Mar. Ecol. Прог. Сер. 471, 73–85. DOI: 10.3354 / meps10036
CrossRef Полный текст | Google Scholar
Heim-Ballew, H., и Olsen, Z. (2019). Влияние солености и температуры на численность скифозойских медуз в западной части Мексиканского залива. Hydrobiologia 827, 247–262. DOI: 10.1007 / s10750-018-3771-0
CrossRef Полный текст | Google Scholar
Хёвар, С., Малей, А., Болдин, Б., и Перселл, Дж. Э. (2018). Сезонные колебания динамики популяции полипов Aurelia aurita in situ с точки зрения моделирования. Mar. Ecol. Прог. Сер. 591, 155–166. DOI: 10.3354 / meps12387
CrossRef Полный текст | Google Scholar
Hubot, N., Lucas, C.H., and Piraino, S. (2017). Экологический контроль бесполого размножения и соматического роста Aurelia spp. (Cnidaria, Scyphozoa) полипы Адриатического моря. PLoS One 12: e0178482. DOI: 10.1371 / journal.pone.0178482
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Исии, Х., Кацукоши, К. (2010). Сезонное и вертикальное распределение полипов Aurelia aurita на пилоне в самой внутренней части Токийского залива. J. Oceanogr. 66, 329–336. DOI: 10.1007 / s10872-010-0029-5
CrossRef Полный текст | Google Scholar
Ларсон Р. Дж. (1991). Диета, выбор добычи и суточный рацион Stomolophus meleagris, сцифомедузы, питающейся фильтром, из северо-восточной части Мексиканского залива. Estuar. Морской берег. Shelf Sci. 32, 511–525. DOI: 10.1016 / 0272-7714 (91)
CrossRef Полный текст | Google Scholar
Лукас, К. Х., Грэм, В. М., и Видмер, К. (2012). Истории жизни медуз: роль полипов в формировании и поддержании популяций скифомедуз. Амстердам: Elsevier Ltd.
Google Scholar
Lutterschmidt, W. I., and Hutchison, V.H. (1997). Критический тепловой максимум: история и критика. Кан. J. Zool. 75, 1561–1574. DOI: 10.1139 / z97-783
CrossRef Полный текст | Google Scholar
Линам, К. П., Гиббонс, М. Дж., Аксельсен, Б. Э., Спаркс, К., Кутзи, Дж. К., Бенджамин, Г. Х. и др. (2006). Медузы настигают рыбу в экосистеме, которая активно вылавливается. Curr. Биол. 16, 1976–1976.
Google Scholar
Мадейра, Д., Нарцисо, Л., Кабрал, Х. Н., и Винагре, К. (2012). Термическая толерантность и потенциальные воздействия изменения климата на прибрежные и устьевые организмы. J. Sea Res. 70, 32–41. DOI: 10.1016 / j.seares.2012.03.002
CrossRef Полный текст | Google Scholar
Malej, A., Kogovšek, T., Ramšak, A., Catenacci, L., Bonnet, D., Carcaillet, F., et al. (2012). Цветение и динамика численности лунных медуз в северной Адриатике. CBM Cah. Биол. Мар. 900: 30, 53: 337. DOI: 10.1016 / j.scitotenv.2018.01.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маркес, Р., Канту, М., Сориано, С., Молинеро, К. Дж., И Боннет, Д. (2015). Картирование распространения и местообитаний Aurelia sp. полипы в лагуне Тау, северо-запад Средиземного моря (Франция). Mar. Biol. 162, 1441–1449. DOI: 10.1007 / s00227-015-2680-2
CrossRef Полный текст | Google Scholar
Маркес, Р., Дарнауд, А. М., Скьярити, А., Тремблей, Ю., Молинеро, К. Дж., Сориано, С. и др. (2019). Динамика и бесполое размножение медузы Aurelia coerulea на стадии бентосной жизни в лагуне Тау (северо-запад Средиземноморья). Mar. Biol. 166: 74.
Google Scholar
Миллер, М.-Э. К., и Грэм, В. М. (2012). Экологические доказательства того, что сезонная гипоксия увеличивает выживаемость и успешность полипов медуз в северной части Мексиканского залива. Дж.Exp. Mar. Biol. Ecol. 432, 113–120. DOI: 10.1016 / j.jembe.2012.07.015
CrossRef Полный текст | Google Scholar
Мюлинг, Б.А., Ли, С.-К., Ламкин, Дж. Т., и Лю, Ю. (2011). Прогнозирование последствий изменения климата для нерестилищ синего тунца (Thunnus thynnus) в Мексиканском заливе. ICES J. Mar. Sci. 68, 1051–1062. DOI: 10.1093 / icesjms / fsr008
CrossRef Полный текст | Google Scholar
Нагата Р. М., Хаддад М. А. и Ногейра М.Младший (2009). Неприятность медуз (Cnidaria, Medusozoa) для креветочных тралов в центральной части южной части Бразильского залива с точки зрения рыбаков-кустарей. Pan Am. J. Aquat. Sci. 4, 312–325. DOI: 10.1371 / journal.pone.0195352
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пек, Л.С., Кларк, М.С., Морли, С.А., Мэсси, А., и Россетти, Х. (2009). Пределы температуры животных и экологическая значимость: влияние размера, активности и скорости изменений. Функц. Ecol. 23, 248–256. DOI: 10.1111 / j.1365-2435.2008.01537.x
CrossRef Полный текст | Google Scholar
Пираино, С., Альери, Г., Мартелл, Л., Маццольди, К., Мелли, В., Милисенда, Г. и др. (2014). Pelagia benovici sp. nov. (Cnidaria, Scyphozoa): новая медуза в Средиземном море. Zootaxa 3794, 455–468. DOI: 10.11646 / zootaxa.3794.3.7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Питт, К.А. и Кингсфорд М. Дж. (2003). Временные и пространственные вариации пополнения и роста медуз медуз, Catostylus mosaicus (Scyphozoa: Rhizostomeae). мар. Freshw. Res. 54, 117–125.
Google Scholar
Пёртнер, Х.-О. (2002). Вариации климата и физиологические основы температурозависимой биогеографии: системная молекулярная иерархия термостойкости у животных. Сост. Biochem. Physiol. Мол. Интегр. Physiol. 132, 739–761.DOI: 10.1016 / s1095-6433 (02) 00045-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перселл, Дж. Э. (2005). Влияние климата на формирование цветков медуз и гребневиков: обзор. J. Mar. Biol. Доц. UK 85, 461–476. DOI: 10.1017 / s0025315405011409
CrossRef Полный текст | Google Scholar
Перселл, Дж. Э. (2007). Влияние окружающей среды на скорость бесполого размножения скифозов Aurelia labiata . мар.Ecol. Прог. Сер. 348, 183–196. DOI: 10.3354 / meps07056
CrossRef Полный текст | Google Scholar
Перселл, Дж. Э. (2011). Цветение медуз и гребневиков совпадает с размножением людей и нарушениями окружающей среды. Annu. Преподобный Mar. Sci. 4, 209–235. DOI: 10.1146 / annurev-marine-120709-142751
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перселл, Дж. Э., Бакстер, Э. Дж., И Фуэнтес, В. (2013). Медузы как продукты и проблемы аквакультуры.Достижения в технологии инкубатория аквакультуры. Амстердам: Эльзевир, 404–430.
Google Scholar
Перселл, Дж. Э., Гувер, Р. А., и Шварк, Н. Т. (2009). Межгодовая изменчивость стробиляции скифозоида Amelia labiata в зависимости от плотности полипов, температуры, солености и условий освещения in situ. Mar. Ecol. Прог. Сер. 375, 139–149. DOI: 10.3354 / meps07785
CrossRef Полный текст | Google Scholar
Перселл, Дж. Э., Малей, А., и Бенович А. (1999a). Возможные связи медуз с эвтрофикацией и рыболовством. Побережье. Эстуар. Stud. 55, 241–263. DOI: 10.1029 / ce055p0241
CrossRef Полный текст | Google Scholar
Перселл, Дж. Э. и Стердевант, М. В. (2000). Выбор добычи и диета пересекаются между зоопланктивными медузами и молодыми рыбами в проливе Принца Уильяма, Аляска. Mar. Ecol. Прог. Сер. 210, 67–83. DOI: 10.3354 / meps210067
CrossRef Полный текст | Google Scholar
Перселл, Дж.E., Uye, S. I., and Lo, W. T. (2007). Антропогенные причины цветения медуз и их прямые последствия для человека: обзор. Mar. Ecol. Прог. Сер. 350, 153–174. DOI: 10.3354 / meps07093
CrossRef Полный текст | Google Scholar
Перселл, Дж. Э., Уайт, Дж. Р., Немази, Д. А., и Райт, Д. А. (1999b). Влияние температуры, солености и питания на бесполое размножение и численность сцифозного Chrysaora quinquecirrha . Mar. Ecol. Прог.Сер. 180, 187–196. DOI: 10.3354 / meps180187
CrossRef Полный текст | Google Scholar
Киньонес, Дж., Монрой, А., Ача, Э. М., и Мианзан, Х. (2013). Прилов медуз снижает прибыль при промысле анчоусов у побережья Перу. Рыба. Res. 139, 47–50. DOI: 10.1016 / j.fishres.2012.04.014
CrossRef Полный текст | Google Scholar
Риаскос, Дж. М., Виллегас, В., и Пачеко, А. С. (2014). Состав рациона большой сцифозойной медузы Chrysaora plocamia в высокопродуктивном центре апвеллинга на севере Чили. Mar. Biol. Res. 10, 791–798. DOI: 10.1080 / 17451000.2013.863353
CrossRef Полный текст | Google Scholar
Риппингейл Р. и Келли С. (1995). Размножение и выживаемость Phyllorhiza punctata (Cnidaria: Rhizostomeae) в сезонно меняющемся режиме солености в Западной Австралии. мар. Freshw. Res. 46, 1145–1151.
Google Scholar
Робинсон, К. Л., и Грэм, В. М. (2013). Многолетнее изменение численности северной части Мексиканского залива scyphomedusae Chrysaora sp и Aurelia spp.со ссылками на изменчивость климата. Лимнол. Oceanogr. 58, 235–253. DOI: 10.4319 / lo.2013.58.1.0235
CrossRef Полный текст | Google Scholar
Скоррано, С., Альери, Г., Боеро, Ф., Доусон, М. Н., и Пираино, С. (2017). Разоблачение видов Aurelia в Средиземном море: комплексный морфометрический и молекулярный подход. Zool. Дж. Линн. Soc. 180, 243–267.
Google Scholar
Сегура-Пуэртас, Л., Селис, Л., и Кьяверано, Л.(2009). «Medusozoans (Cnidaria: Cubozoa, Scyphozoa и Hydrozoa) Мексиканского залива» в Мексиканском заливе — происхождение, вода и биота , ред. Дж. Туннель, Д. Фелдер и С. Аэрле (Техас, США) : Texas A&M University Press), 369–379.
Google Scholar
Сёдзи, Дж., Кудо, Т., Такацудзи, Х., Кавагути, О., и Касаи, А. (2010). Распространение лунной медузы Aurelia aurita в связи с летней гипоксией в заливе Хиросима, Внутреннее море Сето. Estuar.Морской берег. Shelf Sci. 86, 485–490. DOI: 10.1016 / j.ecss.2009.03.001
CrossRef Полный текст | Google Scholar
Сомеро, Г. Н. (2010). Физиология изменения климата: как потенциал акклиматизации и генетической адаптации будет определять «победителей» и «проигравших». J. Exp. Биол. 213, 912–920. DOI: 10.1242 / jeb.037473
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Спангенберг, Д. Б. (1964). Новые наблюдения за Aurelia . Пер. Являюсь. Microsc. Soc. 83, 448–455.
Google Scholar
Томас, К. Д. (2010). Климат, изменение климата и границы ареала. Дайверы. Дистриб. 16, 488–495. DOI: 10.1111 / j.1472-4642.2010.00642.x
CrossRef Полный текст | Google Scholar
Тернер, Р. Э., Рабалайс, Н. Н., и Джустич, Д. (2017). Тенденции летних температур придонной воды на континентальном шельфе северной части Мексиканского залива с 1985 по 2015 год. PLoS One 12: e0184350.DOI: 10.1371 / journal.pone.0184350
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Vinagre, C., Dias, M., Cereja, R., Abreu-Afonso, F., Flores, A.A., and Mendonça, V. (2019). Верхние тепловые пределы и пределы безопасности по потеплению прибрежных морских видов — исходный уровень индикатора для использования в будущем. Ecol. Инд. 102, 644–649. DOI: 10.1016 / j.ecolind.2019.03.030
CrossRef Полный текст | Google Scholar
Vinagre, C., Leal, I., Mendonca, V., Мадейра, Д., Нарцисо, Л., Диниз, М. С. и др. (2016). Уязвимость к потеплению климата и способность к акклиматизации организмов тропических и умеренных прибрежных зон. Ecol. индика 62, 317–327. DOI: 10.1016 / j.ecolind.2015.11.010
CrossRef Полный текст | Google Scholar
Vinagre, C., Mendonça, V., Cereja, R., Abreu-Afonso, F., Dias, M., Mizrahi, D., et al. (2018). Экологические ловушки на мелководье прибрежных вод — потенциальное воздействие волн тепла на организмы тропических и умеренных зон. PLoS One 13: e0192700. DOI: 10.1371 / journal.pone.0192700
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Willcox, S., Moltschaniwskyj, N.A., and Crawford, C. (2007). Бесполое размножение в сцифистомах Aurelia sp: влияние температуры и солености в экспериментальном исследовании. J. Exp. Mar. Biol. Ecol. 353, 107–114. DOI: 10.1016 / j.jembe.2007.09.006
CrossRef Полный текст | Google Scholar
Долгосрочная экологическая устойчивость неместных тихоокеанских устриц к ожидаемым современным условиям изменения климата
Долгосрочная экологическая устойчивость некоренных тихоокеанских устриц к ожидаемым современным условиям изменения климата
Опубликовано 7 декабря 2020
Наука
Теги: биологический ответ, лаборатория, мезокосм, моллюски, морфология, множественные факторы, Северная Атлантика, физиология, дыхание, соленость, температура
Основные
- Долгосрочные последствия изменения климата для неместных видов редко изучаются
- Тихоокеанские устрицы подверглись потеплению, закислению океана и пониженной солености
- Прогнозируемое потепление и закисление океана на 2100 год не повлияло на приспособленность
- Низкая соленость снижает скорость выведения и увеличивает потребление кислорода
- Долгосрочные наблюдения выявили потенциальные сезонные тенденции физиологических показателей
Реферат
Текущее глобальное перераспределение биоты часто объясняется двумя основными факторами: современным изменением климата (CCC) и неместными видами (NIS).Однако, несмотря на свидетельства синергетического эффекта, исследования, оценивающие долгосрочное влияние условий CCC на приспособленность NIS, остаются редкими. Мы изучили интерактивное воздействие потепления, закисления океана и снижения солености на глобально распространенную морскую шекель Magallana gigas (тихоокеанская устрица) в течение десяти месяцев. Скорость роста, клиренса и потребления кислорода измерялась ежемесячно для оценки индивидуальной пригодности. Более низкая соленость оказала значительный и постоянный эффект на M. gigas , уменьшая и увеличивая уровень клиренса и потребления кислорода, соответственно.Ни прогнозируемое повышение температуры морской воды, ни снижение pH не оказали долгосрочного физиологического эффекта, что указывает на то, что условия, прогнозируемые на 2100 год, не повлияют на физиологию и выживаемость взрослых. Эти результаты предполагают, что M. gigas останется успешной во всем мире ННГ, и прогнозируется, что CCC продолжит способствовать их конкурентному доминированию в ближайшем будущем.
Пак К. Э., Риус М. и Мешковска Н., в печати. Долгосрочная экологическая устойчивость некоренных тихоокеанских устриц к ожидаемым современным условиям изменения климата. Исследования морской среды . Статья (требуется подписка).
Окружающая среда способствует толерантности к наркотикам — ScienceDaily
КОЛЛЕДЖ — Толерантность — главный аспект наркозависимости, и один психолог из Техасского университета A&M считает, что условия, в которых принимаются наркотики, могут быть столь же важным фактором развития толерантности, как и сами наркотики.
Более десяти лет техасский психолог A&M Антонио Сепеда-Бенито изучал, как обучение способствует развитию толерантности, и, по его словам, среда, в которой употребляется наркотик, играет большую роль в его воздействии.
«Если одно и то же количество лекарственного средства вводится в одном контексте, а затем в другом, отличном и отличном контексте, то эффекты препарата различны», — говорит Сепеда-Бенито. «Лекарство имеет гораздо больший эффект в новом контексте, чем в контексте, который связан с введенным лекарством».
Сепеда-Бенито, изучавший эффекты морфина и никотина, называет это явление «усвоенной толерантностью» и говорит, что постепенную десенсибилизацию к лекарству можно развить не только путем многократного использования препарата, но и посредством процесса обучения, который включает признание среда.
Другими словами, человек, принимающий наркотик в условиях, когда он или она обычно употребляет наркотик или даже ожидает его употребления, с меньшей вероятностью ощутит все эффекты наркотика, говорит он. Однако, если тот же человек принимает такое же количество препарата в условиях, где он или она обычно не принимает лекарство, то человек, вероятно, почувствует больший эффект от препарата.
Он говорит, что знакомый контекст может означать любую знакомую среду и / или действия, а также время.Он также отмечает, что чем длиннее интервал между эпизодами, тем больше вероятность развития усвоенной толерантности.
Сепеда-Бенито говорит, что это могло бы объяснить некоторые передозировки наркотиков, при которых человек не принимал большее количество наркотика, а вместо этого принимал наркотик в незнакомой обстановке, что делает термин «передозировка» неправильным.
«Это физиологические изменения, опосредованные психологическими аспектами», — говорит он.
По словам Сепеда-Бенито, подобно мяснику, работающему в морозильнике для мяса, который не страдает от холода во время работы, но может чувствовать себя холодно дома, со временем тело начинает готовиться через обучение к окружающей среде. он используется для употребления наркотиков, что приводит к ослаблению действия препарата.
«Это важно, потому что мы знаем, что толерантность — это симптом лекарственной зависимости, и когда вы становитесь толерантным к наркотику, вам нужно принимать больше лекарства, чтобы получить желаемый эффект», — говорит Сепеда-Бенито. «Чем больше наркотиков вы принимаете, тем более физиологически зависимым вы становитесь».
История Источник:
Материалы предоставлены Техасским университетом A&M . Примечание: содержимое можно редактировать по стилю и длине.
Факторы окружающей среды сыграли ключевую роль в развитии толерантности человека
Отправлено 3 февраля 2021 г.
Давление окружающей среды могло привести к тому, что люди стали более терпимыми и дружелюбными по отношению друг к другу, поскольку необходимость делиться пищей и сырьем стала взаимовыгодной, говорится в новом исследовании.
Исследователи обнаружили, что группы населения, которые совместно используют ресурсы, были более успешными. Предоставлено: Ярослав Шмахель, Pixabay.
Такое поведение не было неизбежным естественным прогрессом, но было обусловлено экологическим давлением, говорится в исследовании Йоркского университета.
Люди обладают замечательной способностью заботиться о людях, не относящихся к их родственникам или местной группе. В то время как большинство других животных склонны защищаться по отношению к представителям других групп, наша естественная терпимость позволяет нам сотрудничать сегодня в глобальном масштабе, как это видно из торговли или международных усилий по оказанию помощи при стихийных бедствиях.
Используя компьютерное моделирование многих тысяч людей, собирающих ресурсы для своей группы и взаимодействующих с людьми из других групп, исследовательская группа попыталась установить, какие ключевые эволюционные факторы могли побудить людей к межгрупповой толерантности.
Суровые условия
Исследование предполагает, что это могло начаться, когда люди начали покидать Африку и в период все более суровой и изменчивой окружающей среды.
Исследование было посвящено периоду от 300 000 до 30 000 лет назад, когда археологические данные указывали на большую мобильность и более частые взаимодействия между различными группами.В частности, это время, когда сырье перемещается на гораздо большие расстояния и между группами.
Исследователи обнаружили, что популяции, которые совместно используют ресурсы, с большей вероятностью будут более успешными и с большей вероятностью выживут в суровых условиях окружающей среды, где происходят вымирания, чем те популяции, которые не живут за границей.
Однако в богатой ресурсами среде совместное использование было менее выгодным, а в чрезвычайно суровых условиях население слишком мало для того, чтобы совместное использование было осуществимо.
Допуск
Пенни Спикинс, профессор археологии происхождения человека в Йоркском университете, сказал: «То, что наше исследование демонстрирует важность терпимости для человеческого успеха, возможно, удивительно, особенно когда мы часто думаем о предыстории как о времени конкуренции, однако мы убедились, что в ситуациях, когда люди с излишками делятся за границу с нуждающимися, каждый выигрывает в долгосрочной перспективе ».
Доктор Дженнифер К. Френч, преподаватель археологии палеолита Ливерпульского университета, добавила: «Результаты нашего исследования также имеют важное значение для более широких дискуссий об увеличении количества примеров инноваций и более высоких темпах культурной эволюции, которые произошли в этот период.